
Elamena nara, Rahayu & Nugroho, 2019
|
publication ID |
https://doi.org/ 10.26107/RBZ-2019-0014 |
|
publication LSID |
lsid:zoobank.org:pub:D1B44EA6-88C3-4A02-88FD-FAFC8A228398 |
|
persistent identifier |
https://treatment.plazi.org/id/3E1F495F-3ED6-4D87-ADA6-7BE2F519F58C |
|
taxon LSID |
lsid:zoobank.org:act:3E1F495F-3ED6-4D87-ADA6-7BE2F519F58C |
|
treatment provided by |
Carolina |
|
scientific name |
Elamena nara |
| status |
sp. nov. |
Elamena nara View in CoL new species
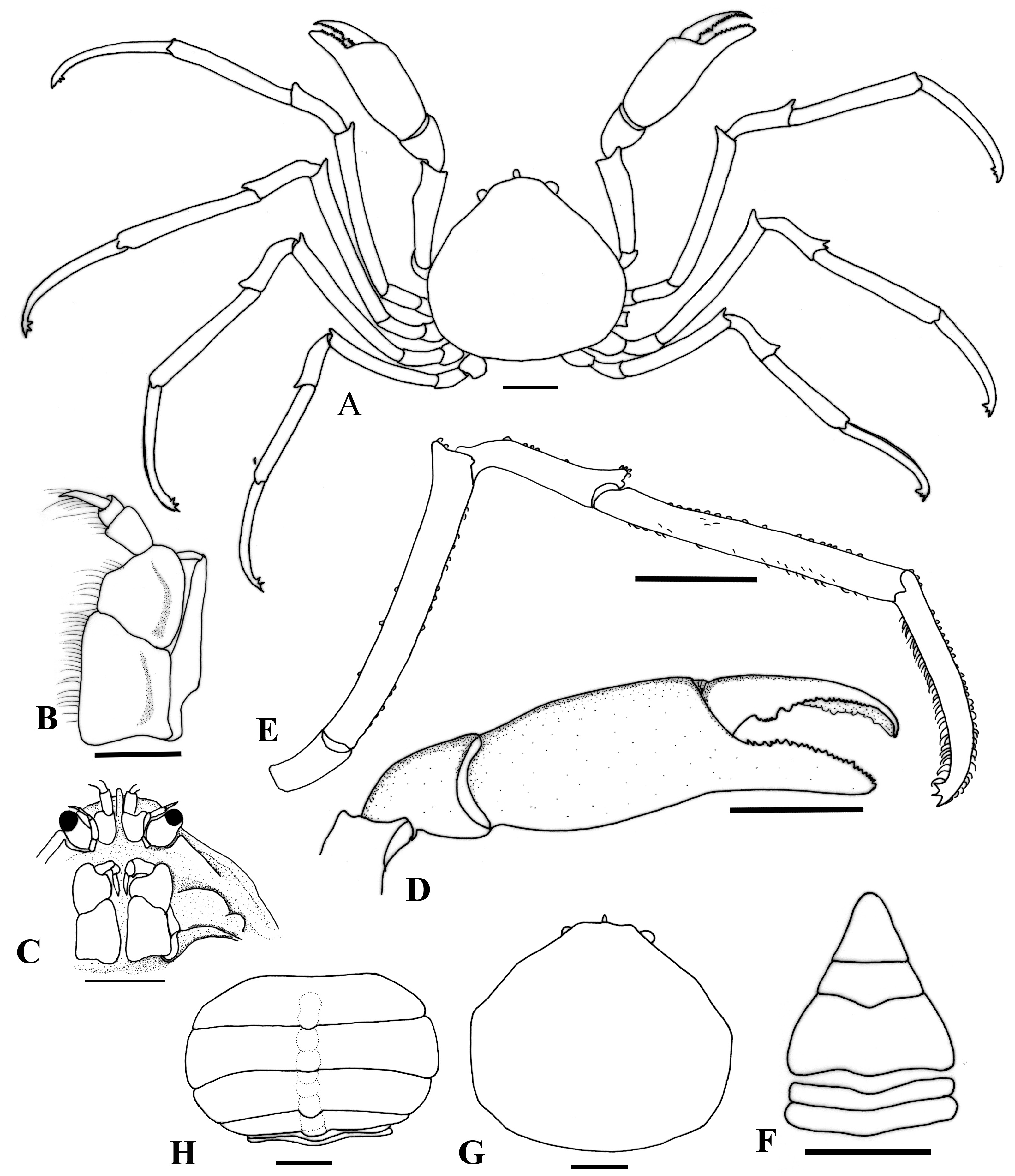
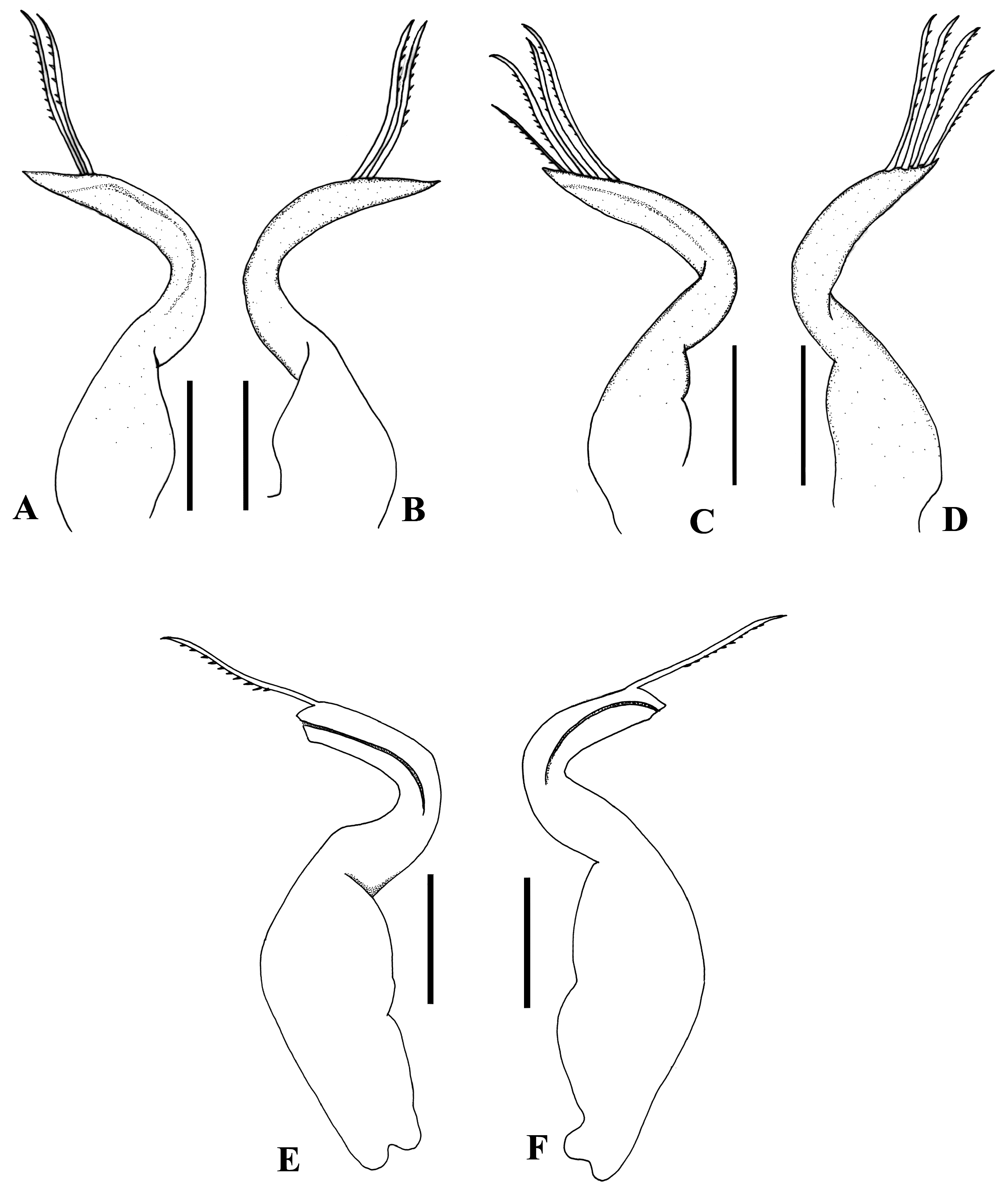
( Figs. 1 View Fig , 2A, B View Fig , 3A View Fig )
Material examined. Holotype, male (3.6 × 3.4 mm) ( MZB Cru 4895), Teluk Nara , Lombok Island, Indonesia, May 2010 . Paratypes, 4 ovigerous females (4.5 × 4.2 mm, 4.1 × 3.6 mm) ( MZB Cru 4896), (5.1 × 4.5 mm, 4.2 × 3.8 mm) ( ZRC2019.0592 View Materials ), Pandanan , Lombok Island, Indonesia, 25 October 2009 .
Description. Carapace ( Fig. 1A, G View Fig ) subpentagonal, slightly broader than long; dorsal surface slightly concave, gastrocardiac groove present but faint. Rostrum semi-truncated, anterior margin slightly convex medially, with sparse bristles, laterally continuous with anterolateral margin of carapace; ventral keel deep dorsoventrally, triangular in lateral view, partially visible from dorsal view, posteriorly continuous with prominent interantennular septum. Postocular tooth distinct, triangular, with rounded tip, not visible from dorsal view. Anterolateral margin slightly sinuous ( Fig. 1A, G View Fig ), slightly concave near base of rostrum; first, second anterolateral teeth replaced by slight convexity, rounded boundary with posterolateral margin, respectively in male, second anterolateral tooth slightly indicated in female; posterolateral margin convex, not well demarcated from posterior margin in male, rounded and well demarcated from posterior margin in female; posterior margin slightly convex. Pterygostomian region with 2 lobes ( Fig. 1C View Fig ), one lobe at opposite ischio-meral suture of third maxilliped, broadly triangular, one much smaller lobe on anterior to base of cheliped coxa. Epistome length subequal to breadth, posterior margin sinuous, with median notch.
Antennules well developed; basal antennular articles placed between bases of eyes, medially separated by strong antenullar septum (proepistome), with relatively short distoexternal protuberance. Antennae directed anteriorly; basal antennal article developed from below base of eye. Eyes well developed, short, stout, partially visible from anterolateral margin in dorsal view.
Third maxillipeds ( Fig. 1B View Fig ) subquadrat, covering most buccal cavity when closed; exopod narrow, distal end not reaching disto-external angle of merus, with distinct flagellum; ischium subquadrate, longer than merus, internal margin straight, longer than external margin; merus subtriangular, with longitudinal ridge along submedian portion of outer surface, internal margin straight, shorter than external margin.
Male thoracic sternum wide, glabrous; sternites 1–3 fused, partially hidden by third maxillipeds, sunken toward buccal cavity; prominent suture at sternite 3/4; sternopleonal cavity moderately deep, anteriorly reaching level of anterior border of sternite 4, press-button situated at lateral slope of sternopleonal cavity near suture of 5/6.
Chelipeds ( Fig. 1A View Fig ) subequal, stout in male, more slender in female; merus rounded in cross-section; carpus rounded, distal inner angle blunt; palm slightly swollen, chela ( Fig. 1D View Fig ) with fingers equal in length, spoon-shaped, no wide gap when closed, cutting edges lined with small teeth interspersed with much smaller teeth, with larger subproximal tooth on dactylus.
P2−P5 ( Fig. 1A, E View Fig ) long, slender, most of surfaces with sparse bristle, extensor and flexor margins lined with short setae except for dactyli which lined with moderately long setae; P3 longest, P5 shortest. Length of meri subequal to propodi; meri, carpi of all legs with distal acute extension on extensor margin; distal one-third of dactyli gently curving inward, terminating in a sharp claw, with 2 subdistal teeth on flexor margin, directed backwards.
Male pleon with 5 somites, including pleotelson ( Fig. 1F View Fig ). Third, fourth pleonal somites fused, gradually narrowed over distal half, with slightly convex lateral margins; lateral margin of fifth somite straight; pleotelson triangular, apex rounded, 1.6 times as long as fifth somite. G1 ( Figs. 2A, B View Fig , 3A View Fig ) stout, strongly curved, forming C-shaped; tip pointed, with 2 subterminal plumose setae.
Female pleon subcircular ( Fig. 1H View Fig ), wide, covering entire thoracic sternum, with six free somites including pleotelson; pleotelson subquadrate, distal margin slightly concave medially, lined with short setae. Vulvae small, placed on medial fused plate of thoracic sternum, at level of lateral end of sternite 4.
Etymology. The new species is named after the type locality Teluk Nara, Lombok Island, Indonesia. The name is used as a noun in apposition.
Type locality. Teluk Nara (Nara bay), northern Lombok Island, Indonesia .
Remarks. Most species included in the truncated rostrum group of the genus Elamena possesses swollen or moderately swollen palm of the chelipeds, thin, slender ambulatory legs (except for E. panglao ), with the meri longer than the propodi (except for E. truncata and E. cristatipes ), and a dome-shaped female pleon covering the sternum ( Gordon, 1940; Lucas, 1980; Ng & Chuang, 1996; Ng & Richer de Forges, 1996; Naruse et al., 2008; Husana et al., 2013; Kulatbong et al., 2013). The length of the meri and propodi of the ambulatory legs of E. nara new species and E. truncata are subequal, but the shape of the carapace, third maxilliped, male pleon, and G1 are different. In E. nara the lateral margins of the carapace is slightly convex without visible teeth in male, hardly visible in females, the ischium of the third maxilliped is as broad as the merus, the margins of the fifth somite of the male pleon is straight, and the G1 have only two setae ( Figs. 1 View Fig , 2A View Fig , 3A View Fig ). In E. truncata the lateral margins of the carapace has a visible angle, the ischium of the third maxilliped is broader than the merus, the margins of the fifth somite of the male pleon is sinuous, and the G1 have nine subterminal setae (cf. Ng & Chuang, 1996: fig. 30). Furthermore, the merus of the cheliped of E. truncata is armed with subdistal strong spine on dorsal margin (cf. Ng & Chuang, 1996: fig. 30D) which is absent in E. nara . Elamena cristatipes with the propodus of ambulatory legs much longer than the merus, the rounded rostrum, and tip of G1 pointed distally, with three subdistal setae (cf. Ng & Chuang, 1996: fig. 7A, D, F, G) separated this species from E. nara . Elamena panglao is clearly different from the new species by proportionally short ambulatory legs and long male pleotelson which is 2.2 times as long as the fifth somite (cf. Naruse et al., 2008: fig. 5a, c, d) versus long and slender ambulatory legs, and relatively short male pleotelson, only 1.6 times as long as the fifth somite in E. nara .
The different shape of the carapace, male pleon and pleotelson, and G1 separated E. nara from E. abrohensis , E. simplidenta , and E. sundaica . In E. abrolhensis the carapace has more pronounced lateral angle, the male pleon is broad with subcircular pleotelson, and the G1 is slender, S-shaped with 12 subdistal setae (cf. Gordon, 1940: figs. 6b, 7a, c); in E. sundaica the carapace has strongly produced, dentiform lateral angle with its anterolateral angle lobiform, the male pleon is broad with subcircular pleotelson, and the G1 is less curved with 11 subdistal setae (cf. Ng & Chuang, 1996: fig. 12A, I−K), while in E. nara the carapace has no visible angle on its anterolateral margins, the male pleon is narrower with blunt triangular pleotelson, and the G1 is stout, C-shaped, with only two subdistal setae ( Figs. 1A, F, G View Fig , 2A View Fig , 3A View Fig ). Elamena simplidenta has been known only by female specimens. Comparison of the females of the two species showed morphological differences. The female carapace of E. simplidenta has a pronounced anterolateral angle (cf. Ng & Chuang, 1996: fig. 11A), but that of E. nara is replaced by slight convexity ( Fig. 1G View Fig ). The shape of female pleon is broader and relatively more rounded in E. nara ( Fig. 1H View Fig ) compare to slightly more oval in. simplidenta (cf. Ng & Chuang, 1996: fig. 11J).
Several morphological differences distinguish E. nara from E. castanea and E. vesca . The male pleotelson of E. nara , is about 1.6 times as long as the fifth somite, with the lateral margins of the fused third and fourth somites slightly convex ( Fig. 1F View Fig ), and male G1 strongly curved forming C-shaped with pointed tip directed ventrally, with two long setae subdistally ( Figs. 2A, B View Fig , 3A View Fig ). In contrast, E. castanea has longer male pleotelson, about 2.1 times as long as the fifth somite, while in E. vesca the pleotelson is as long as the fifth somite, and both species have the lateral margins of the fused third and fourth somites of pleon clearly convex (cf. Naruse et al., 2008: fig. 3d; Ng & Richer de Forges, 1996: fig. 8E). In E. castanea the male G1 is slightly bent and forming broad C-shaped, tip tapering, directed ventrally and has oblique row of six setae subdistally (cf. Naruse et al., 2008: fig. 3e, f), whereas in E. vesca the male G1 is also strongly curved but the tip is pointed upward and has subterminal setae on both side of its apex (8 setae on right and 4 setae on left) (cf. Ng & Richer de Forges, 1996: 275, fig. 8G, H).
The semitruncated rostrum, the presence of two lobes on the pterygostomian region, and the triangular shape of pleotelson allied the new species with E. mathaei . The two species differ in the position of the lobes on the pterygostomian region and the shape of G1. While the larger lobe is positioned opposite ischio-meral suture of the third maxilliped for both species, in E. nara the smaller lobe is on the anterior to base of the cheliped coxa ( Fig. 1C View Fig ), in E. mathaei the smaller lobe is on the antero-external angle of the buccal cavity (cf. Gordon, 1940: 63, fig. 3a). Furthermore, the G1 of E. nara is not twisted nor expanded in the apex, with only two subdistal setae ( Figs. 2A, B View Fig , 3A View Fig ), in E. mathaei the G1 is twisted, expanded near the apex and lined with long dense setae. Elamena samalensis has three lobiform crests on the pterygostomian region, acute pleotelson and S-shaped G1 (cf. Husana et al., 2013: figs. 1C, D, 2), and E. soonthronkitti has ambulatory dactyl laterally flattened, unarmed (cf. Kulabtong et al., 2013: fig. 10), separated those two species from the new species.
Tesch (1918) recorded E. truncata from Ceram and Ambon, but Gordon (1940) and Lucas (1980) identified the specimens as E. abrolhensis , and Ng & Chuang (1996) referred them as E. aff. truncata . However the identity of E. truncata s. str. has been established by Ng & Chuang (1996) with the neotype from Taiwan, and it is clearly different from Tesch’s specimens. The shape and proportion of the carapace, the third maxilliped, the chela of the cheliped, and the long ambulatory legs of E. nara resembles the male of E. truncata sensu Tesch, 1918 , from Ambon (cf. Tesch 1918: fig. 4, 4a). It is possible that E. nara is actually E. truncata sensu Tesch, 1918 . Furthermore, Ng & Chuang (1996) compared Tesch’s (1918) another specimen ( Tesch, 1918: pl. I, fig. 4b, c) with E. sundaica and recognised the difference in the short ambulatory legs. Its carapace shape, short ambulatory legs and invisible eyes from dorsal view may suggest that it is actually E. panglao . In any case, as suggested by Ng & Chuang (1996), reexamination of Tesch’s specimens is required to ascertain their identities.
Diagnostic characters of E. nara and two other new species below are listed in the Discussion.
Habitat and distribution. Intertidal, sandy beach with gravels and boulders near seagrass bed. Lombok Island, Indonesia.

| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.