
Cryptotis oreoryctes, Woodman, 2011
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00754.x |
|
persistent identifier |
https://treatment.plazi.org/id/03C42E42-BA6B-FFD1-4FCB-F8AF94A94B1C |
|
treatment provided by |
Valdenar |
|
scientific name |
Cryptotis oreoryctes |
| status |
sp. nov. |
CRYPTOTIS OREORYCTES SP. NOV. ( FIGS 9F View Figure 9 , 10E View Figure 10 )
Cryptotis parva tropicalis: Choate, 1970: 269 View in CoL . Part; not Cryptotis parvus tropicalis ( Merriam, 1895) View in CoL .
Paratypes (six individuals): All from Alta Verapaz, Guatemala: dried skins, skulls, and partial skeletons of a young male obtained 2.i.2007 ( USNM 569854 About USNM ) and an adult female obtained 3.i.2007 ( USNM 569878 About USNM ) from the type locality; cranium (but not the associated skin or mandibles, which belong to a younger C. merriami View in CoL ) of an old adult of unknown sex ( BMNH 43.9.15.4) and a previously mounted skin with glass eyes ( BMNH 43.9.15.5) and separately numbered skull ( BMNH 43.10.28.7) of a young adult of unknown sex obtained in the mid-19 th century from ‘near Coban’ and provided to BMNH by specimen dealer John Leadbeater; dried skins and skulls of two young adults of unknown sex obtained in the mid-19 th century by Osbert Salvin from ‘Verapaz’ ( BMNH 68.2.10.5) and from ‘near Coban’ ( BMNH 7.1.1.35). See Woodman (2011) for additional details regarding the history of BMNH specimens .
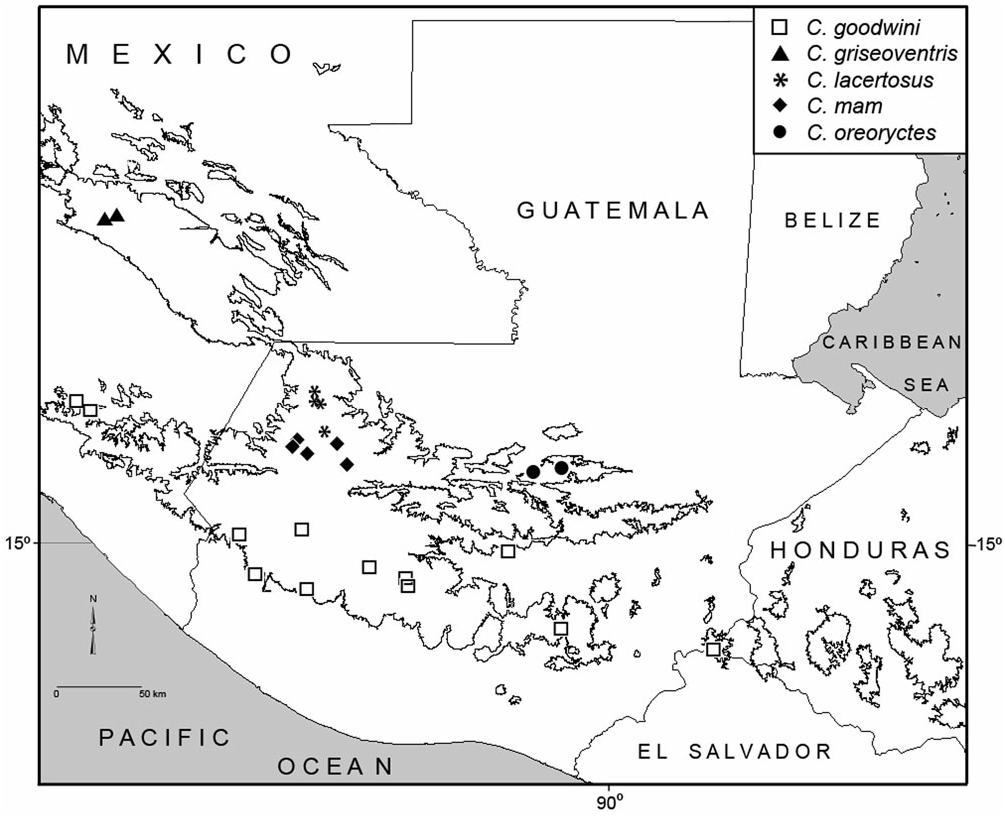
Distribution: Known with certainty only from the vicinity of the type locality ( Fig. 8 View Figure 8 ). The species is probably restricted to remaining forest patches at higher elevations throughout the Sierra de Yalijux. Nineteenth-century specimens in the BMNH from ‘Coban’ may have been taken in forests at higher elevations to the east of that city, Cobán being the closest contemporary population centre of any size.
Etymology: The species name oreoryctes is derived from the classical Greek words OROς (‘mountain’) + ORUSSW (‘to dig’) and signifies ‘digging in the mountains’.
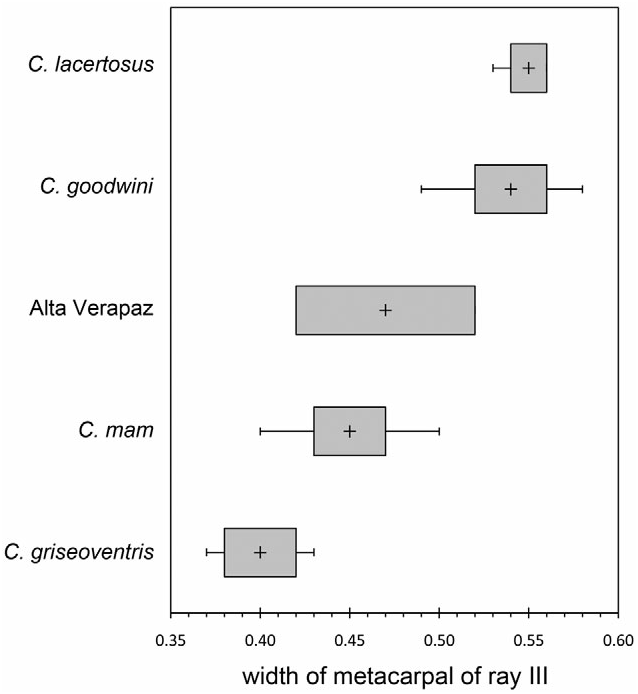
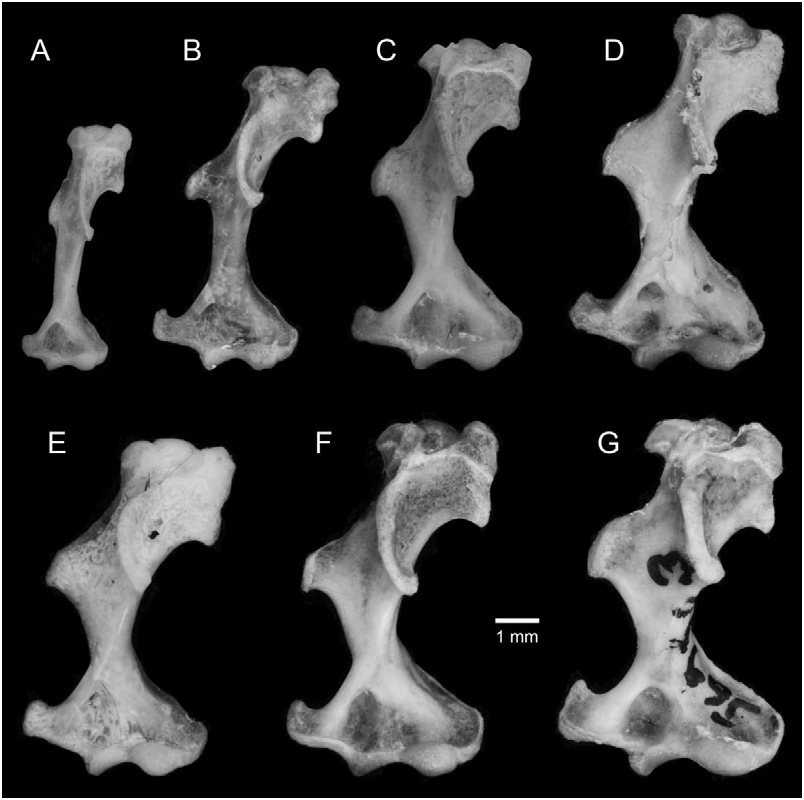
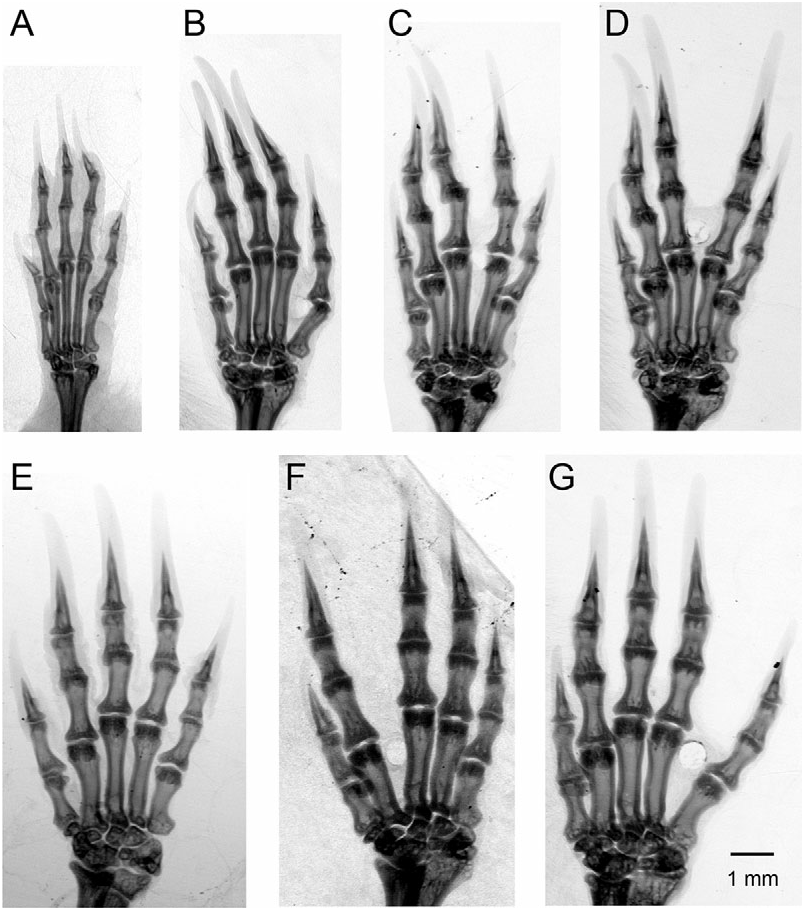
Diagnosis: As a broad-clawed shrew of the C. goldmani group, C. oreoryctes can be distinguished from most other species in the genus by its broad manus and extremely long, broad foreclaws; uncrowded upper unicuspid row in which U 4 is typically aligned and partially visible in labial (fourth unicuspid) view of the rostrum; protoconal basin of M 1 (upper first molar) reduced relative to hypoconal basin; entoconid of M 3 (lower third molar) vestigial or absent; broad humerus with elongated processes ( Fig. 9F View Figure 9 ); short, wide metacarpals and proximal and middle phalanges ( Fig. 10E View Figure 10 ); elongate and broad distal phalanges. Within the C. goldmani group, C. oreoryctes is distinguished by its dark pelage [paler in C. alticola ( Merriam, 1895) , C. goldmani ( Merriam, 1895) , C. lacertosus , C. magnimanus , C. mam , C. peregrinus ( Merriam, 1895) ]; large external body size (larger than C. goldmani , C. griseoventris , C. mam , C. peregrinus ); longer tail (longer than in C. alticola , C. magnimanus ); long skull (longer than in C. alticola , C. goldmani , C. griseoventris , C. magnimanus , C. mam , C. peregrinus ); relatively long unicuspid row (longer than in C. alticola , C. goldmani , C. lacertosus , C. magnimanus , C. mam , C. peregrinus ; but shorter than in C. griseoventris ); moderately broad palate (relatively broader than in C. griseoventris but relatively narrower than in C. alticola , C. goodwini , C. lacertosus , C. magnimanus ); no foramen of the sinus canal (present in C. goldmani , C. peregrinus ); distinct foramen present on tympanic process of one or both petromastoids (absent in C. alticola , C. goldmani , C. goodwini , C. griseoventris , C. lacertosus , C. magnimanus , C. mam , C. peregrinus ); relatively low coronoid process of mandible (lower than in C. goldmani , C. goodwini , C. magnimanus , C. peregrinus ); vestigial entoconid of M 3 sometimes present (always absent in C. griseoventris , C. mam , C. lacertosus ); broad metacarpal and phalanges of ray III of forefoot (broader than in C. griseoventris , C. mam , C. peregrinus , but narrower than in C. goodwini , C. lacertosus ; Figs 6 View Figure 6 , 10 View Figure 10 ); long metacarpal of ray III (longer than in C. goldmani , C. goodwini , C. griseoventris , C. lacertosus , C. mam ); long, broad foreclaws (longer and broader than in C. griseoventris , C. mam , C. peregrinus ); long, broad humerus with prominent teres tubercle and medial and lateral epicondyles (longer and broader than in C. alticola , C. mam , C. peregrinus ; longer than in C. goldmani , but not as obviously massive as C. lacertosus ).
Description: A large member of the genus Cryptotis with a relatively short tail, averaging 30 mm, or 36 ± 3% of head-and-body length ( Tables 1, 10); forefeet broad, foreclaws long and broad. Dorsal pelage approximates Fuscous Black to Chaetura Black; dorsal guard hairs typically 5–6 mm long, with individual hairs up to 7 mm, and indistinctly two-banded: basal five-sixths of hairs silvery grey, grading to a dark brown tip. Ventral pelage paler than dorsum: Mummy Brown to Van Dyke Brown to Sepia. Rostrum long (PL/CBL = 44.5 ± 0.2%; Table 10); postorbital area broad (PO/CBL = 25.9 ± 0.7%); often two obvious dorsal foramina (67%) on the frontals; no ventral extension of the sinus canal or associated foramen posterior to dorsal mandibular articular facet of the skull; typically an obvious foramen dorsal to the dorsal mandibular articular facet present on one or both sides of the skull (100%); typically a distinct foramen on the posteromedial edge of tympanic process of one or both petromastoids [100%; this foramen is much smaller than in C. colombiana Woodman & Timm, 1993 , or C. thomasi ( Merriam, 1897b) – see Woodman & Timm, 1999]; zygomatic plate highly variable in this species, but more typically relatively short (ZP/PL = 21.7 ± 2.9%); anterior border of zygomatic plate typically aligned with mesostyle–metastyle valley or metastyle of M 1, posterior border aligned with posterior edge of M 3 (upper third molar), and at the posterior root of the maxillary process; palate of medium breadth for the genus (M2B/PL = 65.7 ± 1.4%); upper toothrow uncrowded; dentition not bulbous; upper molars lightly to moderately pigmented: medium red to dark red on tips of cones, styles, and cristae; pale to medium red pigment often extends into protoconal basins (67%), and paler red into hypoconal basins, of M 1 and M 2 (upper second molar); unicuspid toothrow relatively long (UTR/ CBL = 13.6 ± 0.1%); posteroventral borders of unicuspids variable, concave to convex; U 4 aligned with the unicuspid row and typically visible in labial view of the rostrum; P 4 (upper fourth premolar), M 1, and M 2 slightly recessed on posterior border; protoconal basin of M 1 reduced relative to hypoconal basin. M 3 small and simple: pigmented parastyle, paracrista, and paracone; reduced precentrocrista and mesostyle may or may not be pigmented; often a short postcentrocrista present; reduced protocone may or may not be pigmented; metacone absent; vestigial, unpigmented hypocone occasionally present. Mandible relatively long and of moderate breadth for the genus; coronoid process of mandible intermediate in height (HCP/ ML = 69.7 ± 1.6%); anterior border of the coronoid process of the mandible joining horizontal ramus at a relatively low angle; long distance from the superior tip of articular process to the posterior border of M 3 (AC3/ML = 80.5 ± 4.1%); articular process generally moderately tall and wide, with a moderately broad lower articular facet; inferior sigmoid notch deep; posterior border of lower incisor aligned approximately with posterior border of hypoconid of P 4 (lower fourth premolar); P 3 (lower third premolar) relatively long and low; entoconid of M 3 vestigial or absent. Metacarpals and proximal and middle phalanges relatively short and broad ( Table 2); distal phalanges elongate and broad; humerus considerably shortened and broadened with elongate processes ( Fig. 7 View Figure 7 ) and dorsoventrally elongate head; medial epicondyle of humerus hooked dorsally; posterior edge of falciform process of the tibia with a deep sulcus.
Comparisons
Like most other members of the C. goldmani group, C. oreoryctes can be most easily distinguished from Guatemalan members of the C. parvus group (i.e. C. tropicalis ) and C. nigrescens group (i.e. C. mayensis , C. merriami ) by its much larger body size (mean HB for C. tropicalis = 68 ± 7; C. mayensis = 69 ± 7; C. merriami = 69 ± 4); longer pelage; broader forefeet and longer, broader foreclaws ( Fig. 10 View Figure 10 ); greater skull size (mean CBL for C. tropicalis = 17.8 ± 6; C. mayensis = 19.2 ± 0.5; C. merriami = 19.5 ± 0.4); and broader, more robust humerus with longer processes ( Fig. 9 View Figure 9 ). For additional measurements and characters of those species, see Woodman & Timm (1992, 1993).
Amongst members of the C. goldmani group in Guatemala and Chiapas ( C. goodwini , C. griseoventris , C. lacertosus , C. mam ), C. oreoryctes has approximately the same external body size as C. lacertosus ( Table 1). Externally, the manus is intermediate in size between those of C. mam and C. goodwini , and the foreclaws are within the size range of those of C. goodwini . Ranges of measurements for individual bones of the forefeet overlap those of C. mam , C. lacertosus , and C. goodwini . Proportionally, however, the metacarpals, proximal phalanges, and distal phalanges of C. oreoryctes tend to be relatively long and narrow ( Tables 3–5). The humerus of the new species is similar in size and in the prominence of its processes to that of C. goodwini ( Fig. 9 View Figure 9 ).
Measurements and indices for C. oreoryctes , C. goodwini , C. griseoventris , C. lacertosus , and C. mam are provided in Tables 1 and 10. Measurements for other species to which C. oreoryctes is compared are provided in parentheses. Additional measurements and characters can be found in Woodman & Timm (1999, 2000), Woodman & Morgan (2005), and Woodman (2010).
Cryptotis alticola : Cryptotis oreoryctes averages larger in body size ( C. alticola: HB = 79 ± 5; WT = 11 ± 3 g) and has a longer tail ( C. alticola: TL = 26 ± 2; TL/HB = 33 ± 4%). The forefeet are of similar size, but the foreclaws are broader. Cryptotis oreoryctes averages larger in skull size ( C. alticola: CBL = 20.2 ± 0.5; BB = 10.4 ± 0.2); has a relatively longer rostrum ( C. alticola: PL /CBL = 43.2 ± 1.2%); relatively narrower palate ( C. alticola : M2B/PL = 70.9 ± 3.0%); and a longer unicuspid row ( C. alticola: UTR = 2.5 ± 0.1; UTR/CBL = 12.3 ± 0.4%). Cryptotis oreoryctes lacks a vestigial foramen of the sinus canal (present in 52% of C. alticola ) and tends to have an obvious foramen dorsal to the dorsal articular facet for the mandible (present in 39% of C. alticola ). Its humerus tends to be shorter, but relatively broader with longer processes than that of C. alticola . Cryptotis alticola is endemic to highlands of central Mexico.
Cryptotis goldmani : Cryptotis oreoryctes averages larger in body size ( C. goldmani: HB = 76 ± 5; WT = 8 ± 1). The forefeet are longer and narrower, and the foreclaws are shorter and narrower. Cryptotis oreoryctes averages larger in skull size ( C. goldmani: CBL = 19.6 ± 0.5; BB = 10.2 ± 0.2); has a longer unicuspid row ( C. goldmani: UTR = 2.3 ± 0.1; UTR/ CBL = 11.9 ± 0.6%); longer and broader palate ( C. goldmani: PL = 8.7 ± 0.3; M2B = 5.8 ± 0.1); and relatively low coronoid process ( C. goldmani: HCP / ML = 80.5 ± 2.6%). Cryptotis oreoryctes lacks a foramen of the sinus canal (well developed in C. goldmani ) and typically has an obvious foramen dorsal to the dorsal articular facet (present in 16% of C. goldmani ). Its humerus is longer, and the processes, particularly teres tubercle and medial epicondyle, are proportionately shorter, but broader. The bones of the forefoot average longer and slightly broader; in ray III, however, the metacarpal averages much longer ( C. goldmani: ML = 2.67 ± 0.28) and the middle phalanx much shorter ( C. goldmani: MPL = 1.09 ± 0.27). Cryptotis goldmani is endemic to highlands in the Mexican states of Guerrero and Oaxaca.
Cryptotis goodwini : Cryptotis oreoryctes averages smaller in body size. The manus is smaller overall, and the foreclaws are shorter and much narrower.
Abbreviations of variables as in Table 1.
Statistics are mean ± SD and range.
Cryptotis oreoryctes averages a longer, narrower skull ( Table 1 – CBL, BB, PO); broader zygomatic plate; longer toothrow; relatively short coronoid process of the mandible; and relatively short posterior length of the mandible. The humerus is generally similar to that of C. goodwini ( Table 2; Fig. 9 View Figure 9 ). Ray III of the forefoot averages longer and narrower; the middle and distal phalanges, however, are shorter ( Table 4; Fig. 6 View Figure 6 ). Cryptotis goodwini occurs in highlands from Chiapas, Mexico, to Honduras.
Cryptotis griseoventris : Cryptotis oreoryctes averages larger externally ( Table 1), and the forefeet appear larger, with longer, broader foreclaws. It also averages larger in nearly all craniomandibular measurements; has a relatively broad palate ( Table 10); more likely to have broad unicuspids with a more convex posteroventral surface; and more likely to possess a vestigial entoconid on M 3. The humerus of C. griseoventris is unknown. The bones and claws of the forefoot are longer and broader ( Table 4; Fig. 6 View Figure 6 ). Cryptotis griseoventris is endemic to northern highlands of Chiapas, Mexico.
Cryptotis lacertosus : Cryptotis oreoryctes has distinctly darker pelage. It averages nearly as large in external measurements, but the manus is smaller, and the foreclaws are shorter and narrower. Cryptotis oreoryctes averages nearly as large in skull size ( Table 1), but has a relatively long rostrum ( Table 10); relatively broad palate; longer toothrow; longer mandible; relatively short posterior length of the mandible. It more often has broad unicuspids with a more convex posteroventral surface, and it is more likely to possess a vestigial entoconid on M 3. The humerus of C. oreoryctes is shorter and less massive ( Table 2; Fig. 9 View Figure 9 ). Most metacarpals and phalanges of the forefoot are longer and often narrower; the middle phalanx of ray III, however, is shorter ( Table 4; Fig. 6 View Figure 6 ). Cryptotis lacertosus is endemic to the northern Sierra de los Cuchumatanes of western Guatemala.
Cryptotis magnimanus : Cryptotis oreoryctes has distinctly darker pelage. It averages nearly as large in external measurements, but the tail is shorter. Cryptotis oreoryctes has a longer skull ( Table 1), relatively long rostrum and narrow palate ( Table 10); longer maxillary and unicuspid toothrows; longer mandible and mandibular toothrow. It more often has broad unicuspids with a more convex posteroventral surface. Cryptotis magnimanus is known only from the Cordillera de Montecillos of Honduras.
Cryptotis mam : Cryptotis oreoryctes has distinctly darker pelage and averages larger in head-and-body length ( Table 1), but tends to have a relatively short tail ( Table 10). Its manus is larger externally, and the foreclaws narrower. Cryptotis oreoryctes averages larger in nearly all craniomandibular measurements. It averages a longer rostrum; relatively shorter posterior length of the mandible; broader unicuspids with a more convex posteroventral surface; and it is more likely to possess a vestigial entoconid on M 3. The humerus is larger and more massive, with more prominent teres tubercle and epicondyles ( Table 2; Fig. 9 View Figure 9 ). The bones and claws of the forefoot are longer and broader ( Table 3–5; Fig. 6 View Figure 6 ). Cryptotis mam is endemic to the Sierra de los Cuchumatanes, western Guatemala.
Cryptotis peregrinus : Cryptotis oreoryctes is larger ( C. peregrinus: HB = 72 ± 3), but has a relatively short tail ( C. peregrinus: TL /HB = 42% ± 3); larger forefeet and longer, broader foreclaws. It also averages larger in nearly all craniomandibular measurements ( C. peregrinus: CBL = 19.1 ± 0.4; BB = 9.9 ± 0.1; M2B = 5.5 ± 0.2); and has a relatively long zygomatic plate ( C. peregrinus: ZP /PL = 19.3% ± 1.0); relatively long unicuspid toothrow ( C. peregrinus: UTR /CBL = 12.9% ± 0.3); and relatively low coronoid process of the mandible ( C. peregrinus: HCP /ML = 73.1% ± 1.6). Cryptotis oreoryctes tends to have broader unicuspids with a more convex posteroventral surface; lacks a sinus canal and associated foramen (well developed in 94% of C. peregrinus ); and typically has an obvious foramen dorsal to the dorsal articular facet (foramen present in 13% of C. peregrinus ). The humerus is larger and more massive, with more prominent teres tubercle and epicondyles. Cryptotis peregrinus is endemic to highlands in central and western Oaxaca, Mexico.
Remarks
The holotype and two other individuals of C. oreoryctes were captured on a steep, moist, north-facing slope with abundant downed trees and mosses in a cloud forest dominated by oaks, pines, and firs at Chelemhá Cloud Forest Reserve in the Sierra de Yalijux. This site is in a region in which native vegetation is classified as Subtropical Montane Rain Forest ( Holdridge, 1947; MAGA, 2001). Chance of frost ( Tamasiunas et al., 2002) at higher elevations in the Sierra de Yalijux varies from low (2–10%) to moderate (20–50%).
The Chelemhá cloud forest contains a diverse association of small mammals. Based on captures using a combination of Sherman live traps, Museum Special snap traps, and pit-falls over seven nights (c. 3208 trap-nights), the small mammal community includes at least 15 species. It is dominated numerically by Peromyscus grandis Goodwin, 1932 and secondarily by Oryzomys saturatior Merriam, 1901a . Heteromys desmarestianus Gray, 1868 , Oligoryzomys fulvescens ( Saussure, 1860) , Reithrodontomys mexicanus ( Saussure, 1860) , Scotinomys teguina ( Alston, 1877a) , and Sorex veraepacis veraepacis Alston, 1877b are relatively abundant (ten to 20 captures) members of the community, whereas Marmosa mexicana Merriam, 1897a , Nyctomys sumichrasti ( Saussure, 1860) , Peromyscus aztecus ( Saussure, 1860) , Peromyscus beatae Thomas, 1903 , Reithrodontomys microdon Merriam, 1901b , Reithrodontomys sumichrasti ( Saussure, 1861) , Reithrodontomys tenuirostris Merriam, 1901b , and Cryptotis oreoryctes are less common (<ten captures).
Reproductive biology of C. oreoryctes is poorly known. A female with obviously worn dentition was pregnant with three embryos on 4 January. A younger female captured on the same day was parous, but not pregnant. The testes of a young male with nearly unworn dentition that was captured on 3 January measured 2.5 ¥ 2 mm.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.