
Liolaemus jamesi (Boulenger, 1891)
|
publication ID |
https://doi.org/10.11646/zootaxa.4612.3.12 |
|
DOI |
https://doi.org/10.5281/zenodo.4323959 |
|
persistent identifier |
https://treatment.plazi.org/id/03C4115F-D407-7319-FDBA-B2D2C830FD78 |
|
treatment provided by |
Felipe |
|
scientific name |
Liolaemus jamesi |
| status |
|
examined the Liolaemus jamesi holotype (NHMUK 1946.8.12.39) and 20 additional specimens from the Tarapacá Region ( Appendix I). The characteristics for scalation were taken according to Etheridge (1995), Lobo (2005) and Quinteros et al. (2008). The lateral head scalation was taken from the left side. Historical data on the L. jamesi type locality was taken from published field notes of the Ambrose A. Lane field trip ( Lane & Sclater 1897); the elevation and locality coordinates were estimated using Google Earth .
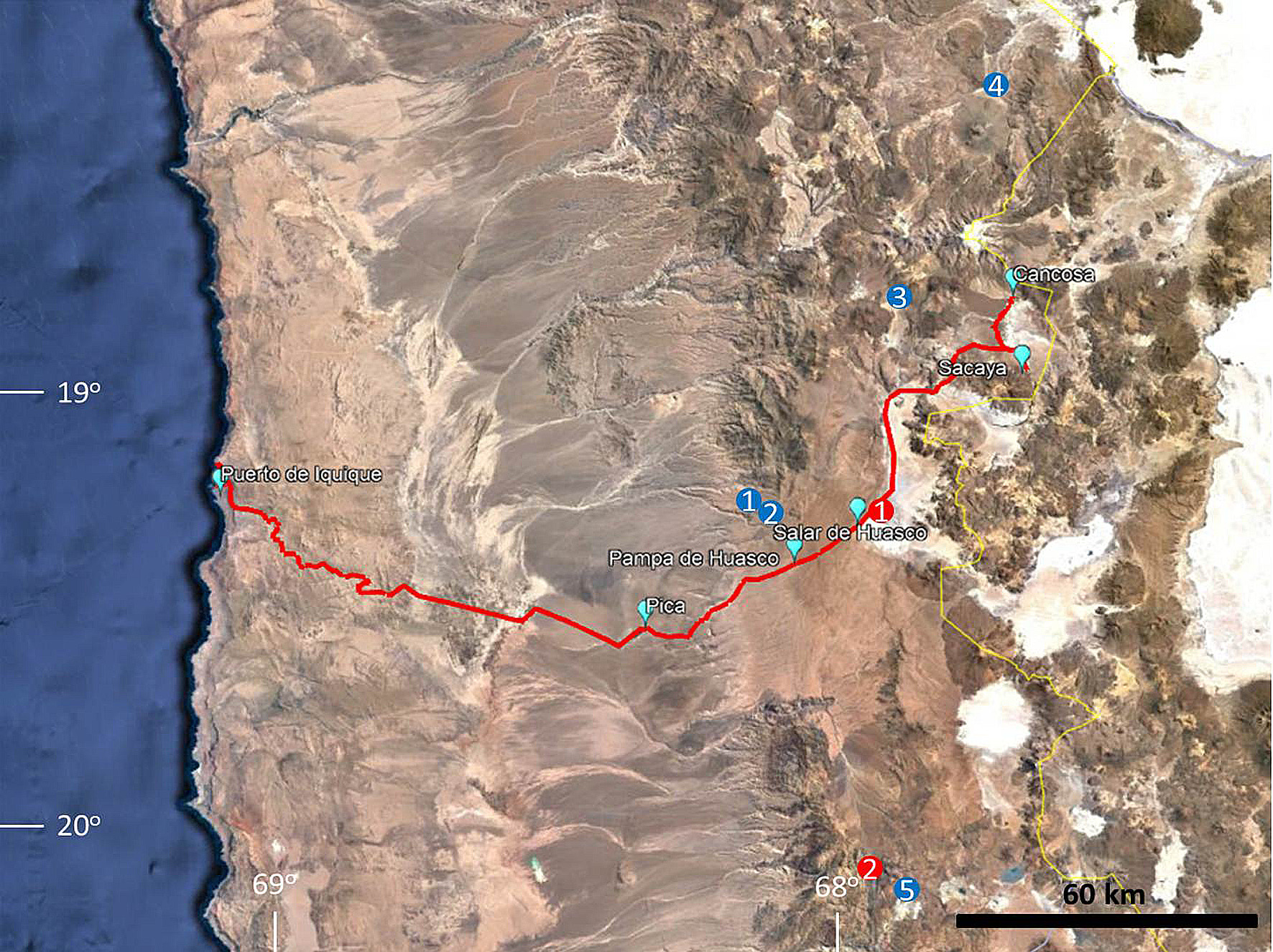
In regards to the type locality, according to Lane and Sclater (1897, p. 10) the journey of Lane in the Tarapacá Province began at Iquique Port GoogleMaps ( 20º13`S – 70º08`W, 30 masl) and then he travelled by the following trajectory outward and by way of return ( Fig. 1 View FIGURE 1 ): Pica ( 20º28`S – 69º18`W, 1300 masl), Pampa de Huasco GoogleMaps ( 20º21`S – 69º01`W, 4000 masl), Laguna (Salar) de Huasco ( 20º17`S – 68º54`W, 3900 masl), Pampa de Sacaya GoogleMaps ( 20º00`S – 68º35`W, 4300 masl) and Cancosa ( 19º51`S – 68º36`W, 3950 m). Neither Boulenger (1891) nor Lane and Sclater (1897) stated the specific location from which the holotype L. jamesi was collected and there had been no new records of this species recorded from this area for more than 100 years.
In fact, the first and only mention of L. jamesi specimens collected from this area is the record from Salar del Huasco ( Moreno et al. 2001), which is at 3900 masl, Boulenger (1891) did however estimate the altitude in which the holotype was collected at 3050-3660 masl. Ruiz De Gamboa and Ortiz (2016) reviewed specimens of L. jamesi from all known distributions, among them one voucher specimen from Altos de Pica ( 20º17`S – 69º05`W, 3545 mals), road to Salar del Huasco, which matched Boulenger`s (1891) altitude range. GoogleMaps Moreover, we collected specimens of L. jamesi (MUAP 122–24, Fig. 2 View FIGURE 2 ) at 17 km W from Laguna del Huasco at 3800 masl ( 20º17`S – 69º03`W). Hence, we are certain that this species occurs in this region at higher elevations GoogleMaps .
We propose a restriction of the type locality of L. jamesi to: road to Salar del Huasco , between 3545 and 3660 masl, Tarapacá Region, Chile, because 1) this transect was explored by the holotype collector Ambrose A. Lane; 2) the other places visited by Lane are out of the elevation range provided by Boulenger (1891); 3) There are records of this species from the Tarapacá Region in Salar del Huasco ( Moreno et al. 2001) and the road to this place ( Ruiz De Gamboa and Ortiz, 2016); and 4) we found specimens at this altitude range matching with the data provided by Boulenger (1891) .
In regards to the holotype ( Fig. 2 View FIGURE 2 ), the measurements and scalation according to Boulenger (1891) are: Adult male. SVL: 100 mm. Tail length (end lost): 90 mm. Head length: 23 mm. Head width: 20 mm. Forelimb length: 47 mm. Hindlimb length: 66 mm. Midbody scales: 52.
The features observed from digital photographs are as follows: Two postrostrals. Two internasals. Hexagonal interparietal scale, with a central, small, and opaque ‘‘parietal eye’’ in the center. The interparietal scale is smaller than the parietals and is surrounded by seven scales. Ten scales between the interparietal scale and the rostral scale. Twenty one scales between the occiput and the rostral. Orbital semicircle incomplete on the right side and formed by 16 scales on the left side; nine supraoculars (enlarged) on right side and seven on the left side; seven supercilliaries. Frontal divided into five scales (1–1–1–2, anterior to posterior). Two scales between the nasal and the canthal. Unfragmented preocular, separated from supralabials by one row of lorilabials. The nasal is separated from the rostral by one scale and is surrounded by seven scales. There is one row of lorilabials between the supralabials and the subocular. There are nine supralabials and seven infralabial scales. The mental shape is irregular. There are five pairs of postmental shields; but the first postmental scale on the right side is not discernible. Temporal scales are juxtaposed, smooth and with some interstitial granules. There are nine temporal scales between the level of the supercilliaries and commissure of the mouth. There are two small scales slightly projected to the auditory meatus, which do not cover it. The auricular scale is not differentiated. There is a well-developed “Y” shaped lateral neck fold, well-developed antehumeral fold and dorso-lateral fold. No gular fold. Dorsal scales are rounded, subimbricate, smooth or slightly keeled, with interstitial granules. The interstitial granules are more abundant on the flanks. Dorsal scales are larger than the ventrals. Dorsal scales: 45, 46 according to Boulenger (1891). Ventral scales are rounded to rhomboidal, smooth, imbricated, with some interstitial granules. Ventral scales: 88. There are six precloacal pores. There is a ventral incision, probably for organ examination, running from the chest to the midpoint of the belly. Suprafemoral and infrafemoral scales are rounded, imbricate, and smooth. Supraantebrachials scales are rounded, imbricate, and smooth. Infraantebrachials scales are rounded, imbricate, and smooth to conical towards the axilla. The dorsal scales of the tail are rounded, subimbricate, weakly keeled, with interstitial granules. The ventral scales of the tail vary from rounded to rhomboidal or triangular, and are imbricate and smooth. Lamellae of fourth finger 23 and lamellae of fourth toe 29 (right hand).
Light brown head, with dark brown spots on the snout, occipital and circumorbital zones. The loreal and temporal zones are light grey. The anterior and middle zones of the subocular scale are grey, but there is a dark brown vertical stripe running from the eigth supralabial to the postocular scale (passing over the posterior zone of the subocular). The dorsal colour is light brown on the right hand side and grey on the left hand side (by deterioration). Some dorsal scales have black spots on the anterior border and there is a black coloured space between the dorsal scales. The keel on the dorsal scales (when it is present) is not black spotted. The flanks are light grey, with a black in between the scales. The dorsal surface of the limbs and the dorsal surface of the tail are both light grey, with black in between the scales. Some scales on the tail (especially in the middle of the tail) is brown shaded in colour. The tip of the tail is brown. The throat, chest and belly are dark brown, with a few light grey scales dispersed. The ventral surface of the hind limbs is light grey in colour with some brown spots. The cloaca is light grey with dark brown in the middle. Precloacal pores are orange. The ventral surface of the tail is light grey with some dark brown spots and a complete dark brown vertebral stripe.
While it is not possible determine the exact type locality, this has also been the case for many other old descriptions of Liolaemus and other South American lizards, the reason being because the descriptions commonly tended to be incomplete or imprecise. Recent publications have been successful in restricting the type locality of some poorly known southernmost South American lizards to bounded geographical ranges, based on the historical data of the field notes, with the goal of producing characterizations and delimitations of the species and their ranges ( Lobo & Etheridge 2013; Quinteros et al. 2008; Troncoso-Palacios & Garin 2013; Troncoso-Palacios et al. 2013; Troncoso-Palacios 2018). In this case, the restriction of the type locality of L. jamesi will allow determination of further morphological and molecular characterization of this taxon, such that it can be compared to species which show some morphologically similarity, especially with the taxa previously considered to be subspecies of L. jamesi or currently considered synonyms of it: L. aymararum Veloso, Sallaberry, Navarro, Iturra, Valencia, Penna & Díaz, 1982 , considered to be a junior synonym of L. jamesi by Pincheira-Donoso and Núñez (2005) and L. jamesi pachecoi Laurent, 1995 , later raised to full species by Langstroth (2011). Moreover, Langstroth (2011), based on the mtDNA data by Schulte and Moreno-Roark (2010), pointed out that mtDNA divergence between L. aymararum and L. pachecoi is enough to recognize both as full species. Furthermore, a recent mt + nuclear DNA phylogeny ( Aguilar-Puntriano et al. 2018) found that L. jamesi is neither the sister species of L. aymararum nor L. pachecoi . We hope that our proposed restriction of the type locality of L. jamesi and the characterization of additional material from its vicinity will facilitate further research on these issues.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.