
Magelona pulchella Mohammed, 1970
|
publication ID |
https://doi.org/10.5281/zenodo.198268 |
|
DOI |
https://doi.org/10.5281/zenodo.6208088 |
|
persistent identifier |
https://treatment.plazi.org/id/03BDDD14-E460-553C-FF41-FDF0BB11FD67 |
|
treatment provided by |
Plazi |
|
scientific name |
Magelona pulchella Mohammed, 1970 |
| status |
|
Magelona pulchella Mohammed, 1970 View in CoL
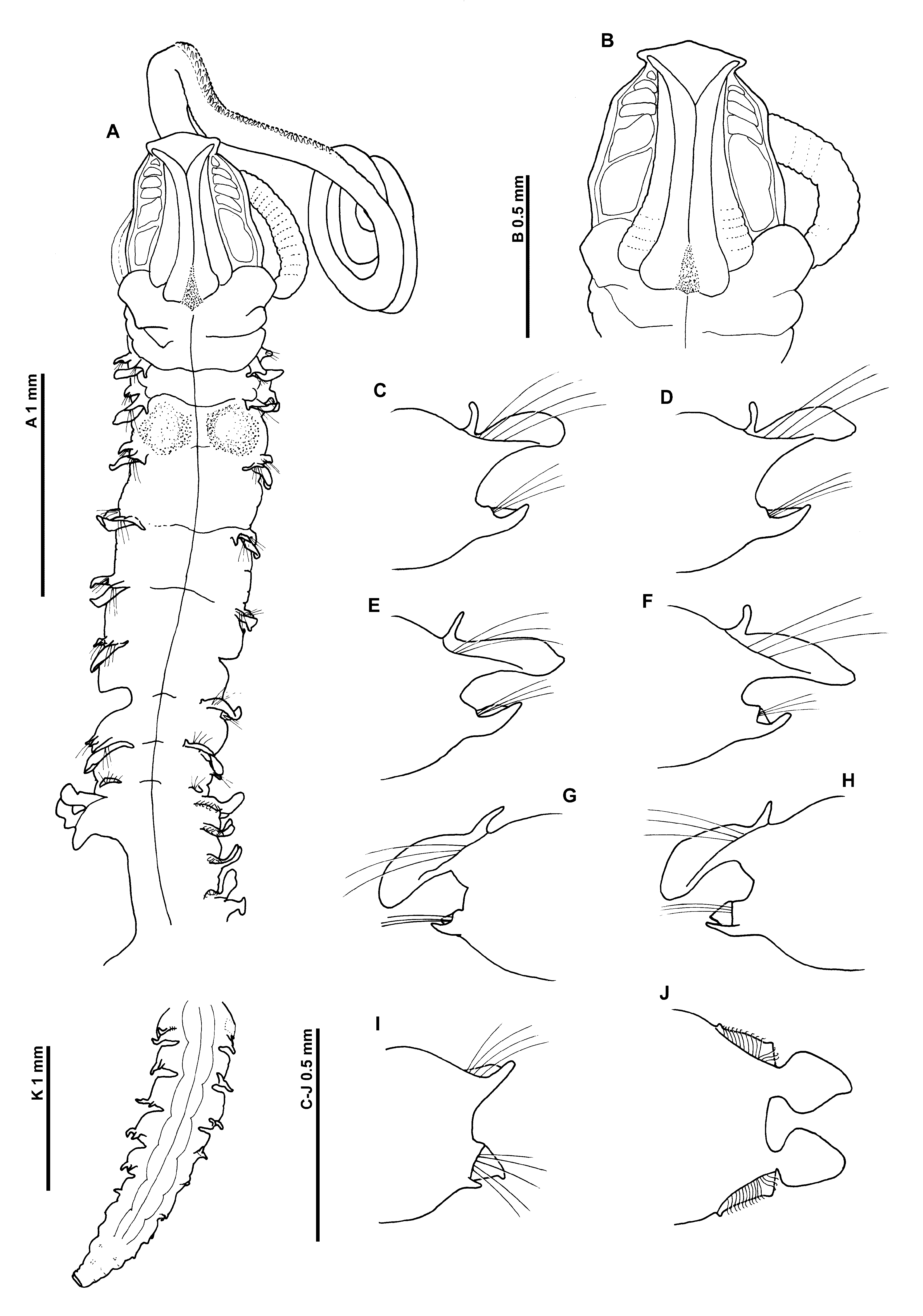
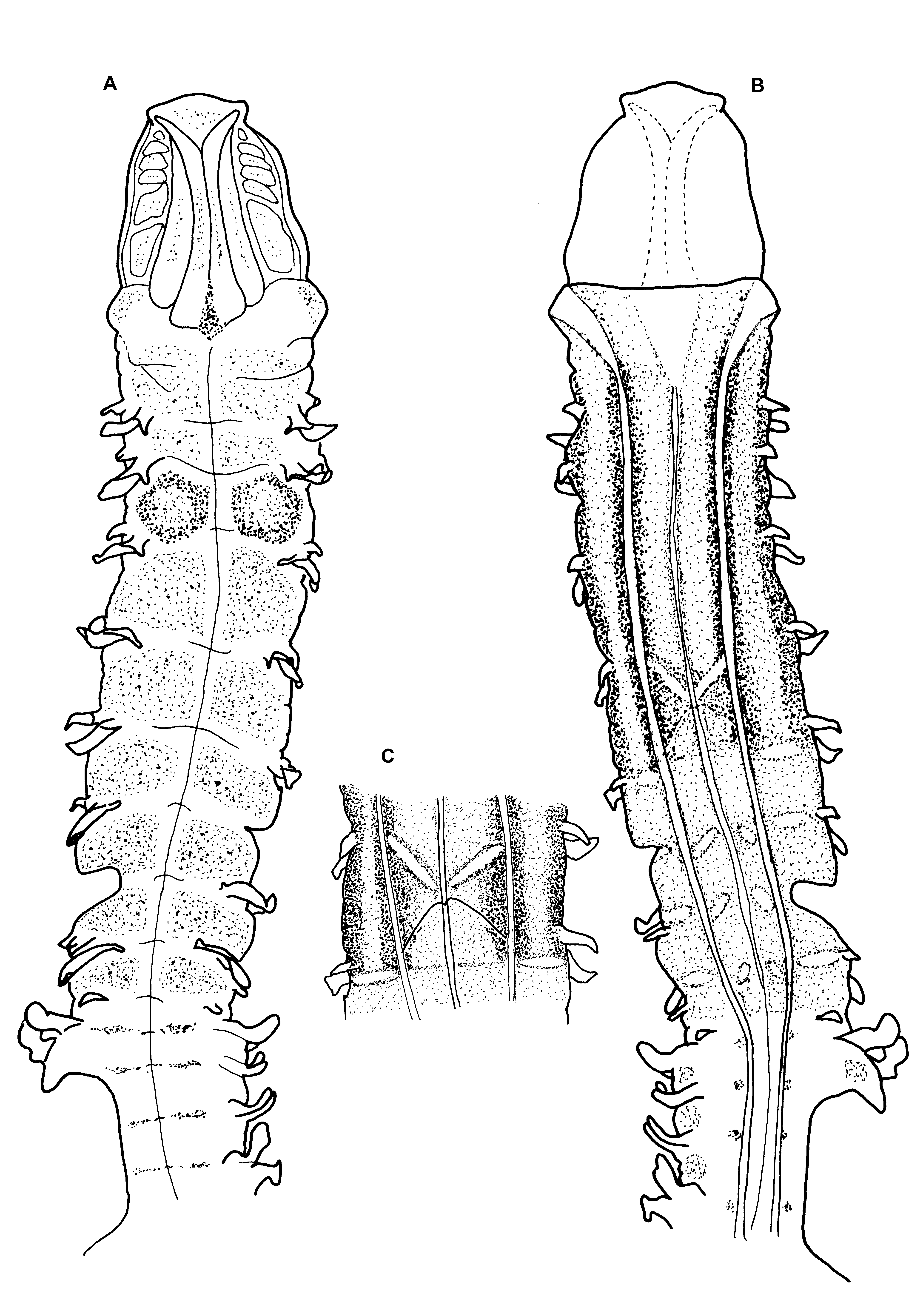
Figures 9–10 View FIGURE 9 View FIGURE 10
Magelona pulchella Mohammed, 1970: 305 View in CoL –306: figs 3–4
Material examined. Kuwait, Arabian Gulf, ( 29°22’N, 48°02’E), in sand, intertidal zone, Holotype ( BMNH 1969.391, c), collected by Murad-B. M. Mohammed, 5/6/1969.
Diagnosis. Prostomium as long as wide, subtriangular, with distinct prostomial horns. Notopodia of chaetigers 1–8 with lanceolate postchaetal lamellae and cirriform dorsal superior processes; neuropodia with digitiform ventral processes, those of chaetigers 7 and 8 with additional subtriangular postchaetal lamellae. Notopodia of chaetiger 9 with broadly rounded low postchaetal lamellae basally encircling chaetae to form digitiform processes beneath chaetae, neuropodia with a subtriangular postchaetal lamellae and small triangular prechaetal processes. All thoracic chaetae capillary. Abdominal lateral lamellae spatulate, bluntly pointed, basally constricted. Hooded hooks bidentate, in 2 groups, vis–à–vis. Posteriorly open pouches present on abdominal chaetigers.
Description. A moderately large species, appearing slightly dorso–laterally flattened; marked difference between abdomen and thorax, thorax narrower and thinner than abdomen ( Fig. 9 View FIGURE 9 A). Dimensions: Prostomium 0.55 mm long, 0.6 mm wide; thorax (including prostomium) 4.0 mm long, 0.5 mm wide; abdomen 0.65 mm wide; total length ~ 36 mm for 108 chaetigers. Note: measurements less than those recorded by Mohammed, although width measurements may have included parapodia (see Mohammed 1973: p.40). Chaetigers 7, 11– 13, 17 (notopodium only) and 24 on the left, and chaetigers 6, 9/10 (neuropodium only) and 29–30 on the right previously dissected. Body weakly attached between chaetigers 64–65.
Prostomium about as long as wide (L:W ratio 0.92), subtriangular, with distinct prostomial horns, anterior margin smooth, triangular; eyes absent. Two pairs of prominent longitudinal dorsal muscular ridges, outer pair shorter, abutting inners for entire length, inner pair abutting for majority of length, diverging distally into corners of frontal horns ( Figs 9 View FIGURE 9 A–B, 10A–B). Distinct large (muscular?) areas either side of ridges varying in shape and size (rounded rectangular, quadrangular or reniform). Proboscis only partially everted, oval; inferiorly longitudinal ridged. Right–hand palp present (left–hand palp detached, present in tube), long; arising ventrolaterally from base of prostomium, fairly robust in comparison to size of specimen, reaching at least chaetiger 25, non–papillated basal region reaching chaetiger 4. Papillae; short proximally, increasing in size, long distally. Initially 3 rows of papillae, with 1 row medially and distally, either side of inconspicuous ventral groove.
Peristomium achaetous, roughly one and a half times size of chaetiger 1. Chaetigers 18 similar; parapodia biramous; notopodia with low triangular prechaetal lamellae confluent with lanceolate postchaetal lamellae, becoming slightly longer throughout thorax ( Figs 9 View FIGURE 9 C–H). Long, slender, tapering cirriform prechaetal superior processes (DML) present on all thoracic chaetigers, except chaetiger 9, all of similar size.
Neuropodia of chaetigers 1–8 with ventral digitiform processes (VNL), in distinctly prechaetal positions, increasing slightly in size to mid–thorax, becoming ventral in position. Small postchaetal expansion, becoming well–developed and triangular on chaetigers 7 and 8.
Chaetiger 9: shorter and narrower than preceding chaetigers ( Fig. 9 View FIGURE 9 A). Notopodial prechaetal lamellae low, expanded as lateral digitiform processes underneath chaetae, confluent with rounded postchaetal lamellae, smaller than on preceding chaetigers ( Fig. 9 View FIGURE 9 I). No superior processes (DML) observed. Neuropodia with triangular postchaetal lamella confluent with low prechaetal ridge, small ventral prechaetal process present (VML of some authors) (right neuropodium unknown due to dissection). Chaetae of all thoracic chaetigers simple winged capillaries (see Mohammed 1970: p.306, figs 4a & b).
Abdominal chaetigers with broad, spatulate lateral lamellae, of about equal size in both rami, basally constricted, bluntly pointed, lamellar shape shows some variation ( Fig. 9 View FIGURE 9 J). Extension of the lateral lamellae behind chaetal rows well–developed, especially in anterior abdomen, apexes triangular. Small triangular processes (DML & VML) present at inner margins of chaetal rows. Posteriorly, lateral lamellae, much reduced, lanceolate.
Abdominal chaetae bidentate hooded hooks of similar size (not drawn to prevent further damage to the specimen). Hooks in two groups, main fangs vis–à–vis, groups at inner margins of chaetal rows with around twice number of chaetae. Initially around 10–14 hooks per rami.
Posteriorly open pouches (C configuration of Fiege et al. 2000 —see discussion) present posteriorly, initially on alternate chaetigers, then more or less on every segment, alternating again towards pygidium. Pouches differing from those seen on M. obockensis and M. heteropoda , less convoluted, simple folds; edges minutely wavy. Pouches observed on 39R, 41L, 43–44L, 43R–85R, 46L–82L, 84L–86L, 87R, 88L, 89R, 90L, 91R, 92L, 93R, 94L, 95R, 96L, 97R, 98L, 99R, 100L. Mohammed stated that the posterior end carries “a short anal cirrus on each side”, these were difficult to discern in the specimen, however two slender anal cirri were observed flattened against the pygidium.
Colour. The colouration originally described by Mohammed was “pinkish white anteriorly and greyish white posteriorly”. Preserved colour uniformly cream/yellow in alcohol, glandular areas noticeable interparapodially in abdomen. Staining with methyl green shows a diffuse overall stain dorsally in the thorax, particularly strong between parapodia of chaetigers 2 and 3 ( Figs 9 View FIGURE 9 A, 10A). This staining is retained long after the majority of stain has dissipated. Additional light speckling on the prostomium and in the anterior abdomen (as light transverse bands). Ventrally the staining is strongest between chaetigers 1 and 5 ( Fig. 10 View FIGURE 10 B), particularly strong either side of the ventral grooves, staining of the posterior thorax is more diffuse. A distinct white X shape present ventrally between chaetigers 4 and 5 ( Fig. 10 View FIGURE 10 C), the upper part being much thicker than lower. Some staining present interparapodially in the abdomen and small speckled patches ventrally either side of the mid–ventral line.
Habitat. Type specimen found in sand, Kuwait, Arabian Gulf. No other records of this species have been found by the author.
Remarks. This redescription details several features either omitted from, or incorrectly reported in the original description. In particular the shape of the prostomium, which in the original drawings is shown with more rounded lateral margins, and which terminates in a somewhat trilobed anterior margin. Further differences are the shape of thoracic neuropodial lamellae (particularly those of chaetigers 7 and 8, which have triangular postchaetal neuropodial lamellae), the presence of a neuropodial ventral process on chaetiger 9, the presence of small processes at the inner margins of chaetal rows on abdominal chaetigers (DML & VML) and abdominal hooded hooks in two groups, vis–à–vis.
Mohammed stated that M. pulchella most resembled M. cornuta (Mohammed 1970: p.306), but differed in not possessing branchiae (cf. Wesenberg-Lund 1949: p.328). Wesenberg-Lund when describing M. cornuta stated “The parapodia are all biramous, those of the first 8 chaetigers with small foliaceous dorsal and ventral cirri or “postsetal lobes”. In front of the dorsal cirrus a small cirriform branchia is found.” The ‘dorsal and ventral cirri’ described by Wesenberg-Lund are likely to be those structures currently termed lateral lamellae, and the ‘small branchia’ the dorsal superior processes (DML), as shown by Mortimer & Mackie (2009).
Magelona pulchella View in CoL shares similarities with M. cornuta View in CoL , M. gemmata Mortimer & Mackie, 2003 View in CoL , M. lenticulata Gallardo, 1968 View in CoL , M. tehuanensis Hernández-Alcántara & Solís-Weiss, 2000 View in CoL , M. berkeleyi Jones, 1971 View in CoL , M. nonatoi Bolívar & Lana, 1986 View in CoL and two unnamed species (J and L) from the Gulf of Mexico ( Uebelacker & Jones 1984) in possessing prostomial horns and in the nature of the thoracic lamellae.
However, it differs from them in possessing bidentate and not tridentate abdominal hooded hooks. It further shares similarities with M. annulata Hartmann-Schröder, 1962 View in CoL , M. crenulifrons View in CoL , M. longicornis Johnson, 1901 View in CoL , M. methae Nateewathana & Hylleberg, 1991 View in CoL , M. pacifica Monro, 1933 View in CoL and one unnamed species (G) from the Gulf of Mexico ( Uebelacker & Jones 1984), all of which possess bidentate hooded hooks. Magelona pulchella View in CoL can be distinguished from M. annulata View in CoL in possessing dorsal superior processes on chaetigers 1–8 and can be distinguished from M. crenulifrons View in CoL and M. methae View in CoL in not having a crenulate anterior prostomial margin. The anterior margin of M. longicornis View in CoL can also be crenulate, however M. longicornis View in CoL further differs from M. pulchella View in CoL in possessing abdominal hooded hooks in one group, main fangs facing laterally. It appears to most closely resemble M. pacifica View in CoL and M. sp. G. but differs in possessing more slender lanceolate postchaetal thoracic lamellae, however, further descriptions of these species would enable better cross– comparisons.
The presence of a characteristic unstained V shape on the venter of M. crenulifrons View in CoL was noted by Mortimer & Mackie (2009; p.190), the point of which was often at the junction between chaetigers 5 and 6. This feature shares many similarities with the unstained X shape present on the venter of M. puchella between chaetigers 4 and 5, particularly the thicker upper region. However, M. pulchella View in CoL is also distinguished from M. crenulifrons View in CoL by deep staining on dorsum between parapodia of chaetigers 2 & 3.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Magelona pulchella Mohammed, 1970
| Mortimer, Kate 2010 |
M. gemmata
| Mortimer & Mackie 2003 |
M. tehuanensis Hernández-Alcántara & Solís-Weiss, 2000
| Hernandez-Alcantara & Solis-Weiss 2000 |
M. methae
| Nateewathana & Hylleberg 1991 |
M. nonatoi Bolívar & Lana, 1986
| Bolivar & Lana 1986 |
M. berkeleyi
| Jones 1971 |
Magelona pulchella
| Mohammed 1970: 305 |
M. lenticulata
| Gallardo 1968 |
M. annulata Hartmann-Schröder, 1962
| Hartmann-Schroder 1962 |
M. pacifica
| Monro 1933 |
M. longicornis
| Johnson 1901 |