
Nisitrus danum Robillard & Tan, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5315.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:56919CD3-9390-4682-855C-0B6026E27A02 |
|
DOI |
https://doi.org/10.5281/zenodo.8132444 |
|
persistent identifier |
https://treatment.plazi.org/id/03BCA678-817F-DB15-FF19-FC56376FFAAC |
|
treatment provided by |
Plazi |
|
scientific name |
Nisitrus danum Robillard & Tan, 2021 |
| status |
|
Nisitrus danum Robillard & Tan, 2021 View in CoL
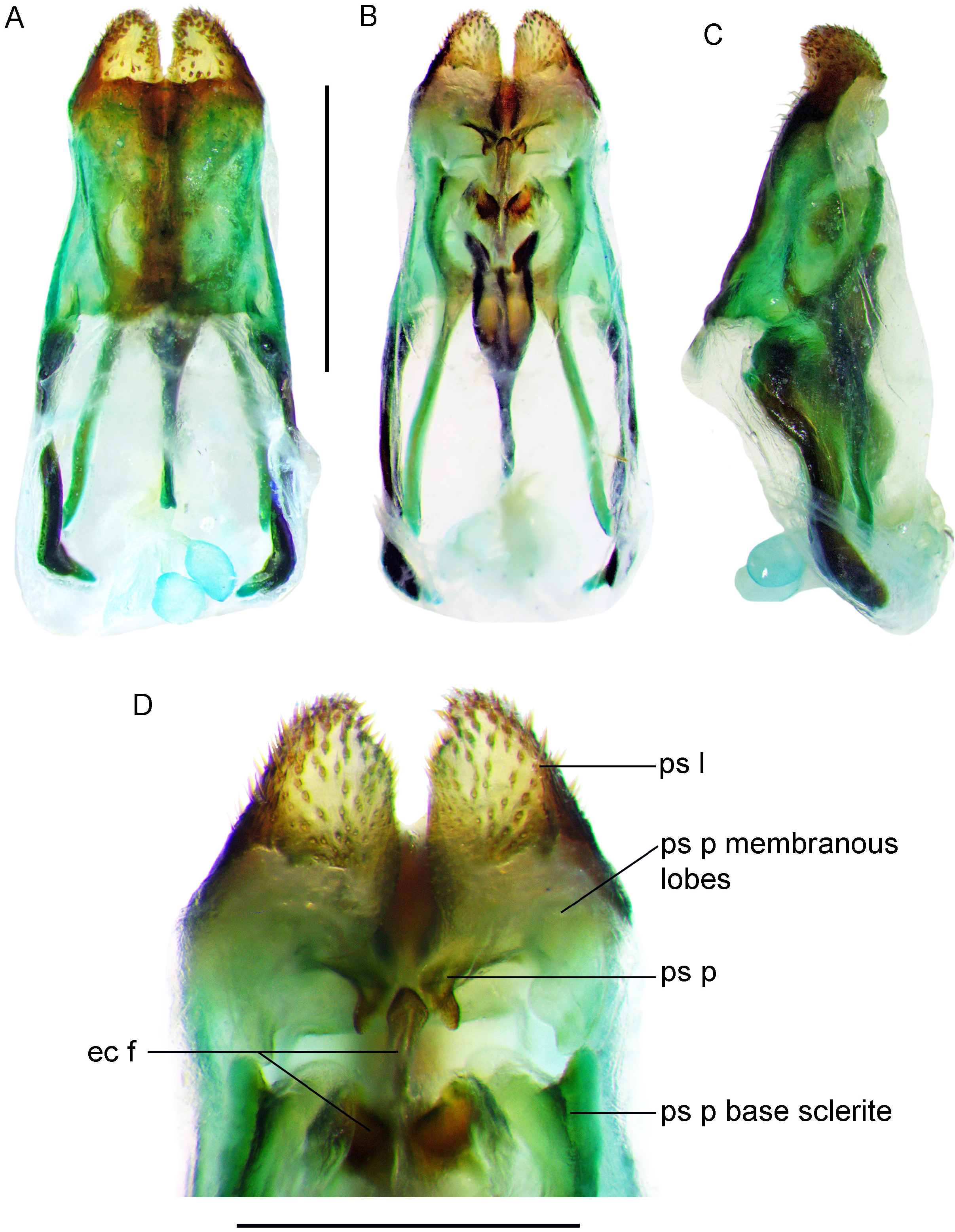
( Figs 1–8 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
Nisitrus danum Robillard & Tan, 2021 , in Tan et al. 2021a: 31
Material examined
MALAYSIA — Sabah • 1J; Kawag Forest Reserve, near rest house; N5.0502 E117.98286, 134.7 m.a.s.l. (GPS SAB22_6); 14.v.2022; afternoon; on plant; call recording MNHN-SO-2023-1478; M.K. Tan & T. Robillard; MNHN-EO-ENSIF11344 • 1♀; Kawag Forest Reserve, near rest house; N5.0502 E117.98286, 134.7 m.a.s.l. (GPS SAB22_ 6); 14.v.2022; afternoon; on plant; molecular sample N67; M.K. Tan & T. Robillard; MNHN-EO-ENSIF11345 • 1♀ (photographed, not collected);Tabin Wildlife Reserve,Lipad mud volcano; N5.20967 E118.50732, 168.9m.a.s.l. (GPS SAB22_11); 16.v.2022; day; T. Robillard observation • 2 juveniles; Tabin Wildlife Reserve; N5.19534 E118.50365, 119.7 m.a.s.l. (GPS SAB22_9); 15.v.2022, night; molecular samples N65–N66; MNHN-EO-ENSIF10978– ENSIF10979 • 1 juvenile (not collected); Sepilok, Rainforest Discovery Centre; N5.87428 E117.93970, 1.x.2019 ( Fig. 3B View FIGURE 3 ) • 1 juvenile; Lahad Datu, N4.962598 E117.801956; 19.viii.2022; twan3253 observation; https://www. inaturalist.org/observations/133043313 • 1♀; Lahad Datu, N4.964485 E117.803471; twan3253 observation; https:// www.inaturalist.org/observations/133303271 • 1♀; Lahad Datu, N4.965797 E117.800246; twan3253 observation; https://www.inaturalist.org/observations/133431471
Additional description
Habitus as shown in Fig. 1A View FIGURE 1 . Face, lateral lobes of pronotum and wide lateral band on FWs vivid yellow in living specimens ( Figs 1A, 1B View FIGURE 1 ). Antennae with 2 main white rings, including 3–6 antennomeres and 2–3 thinner rings.
Male. Metanotal gland well developed, as shown in Fig. 1C View FIGURE 1 . FWs ( Figs 1D, 1E View FIGURE 1 ) dorsal field anterior part black (cells and veins); harp and mirror area infuriated, with black veins; apical field with black cells and yellow veins. Harp with two straight oblique veins. Mirror large, not rounded, separated by one transverse vein. Vein c1 wide. Lateral field with a wide vivid yellow band including veins M, R and Sc, and bases of veins bifurcating from Sc; ventral part of lateral field shiny black, as in females.
Male genitalia ( Fig. 2 View FIGURE 2 ) typical of genus, including ectophallic fold, endophallic sclerite, ectophallic arc and rami, but with pseudepiphallic lophi triangular and relatively stout in dorsal and profile views, narrowly spaced between pseudepiphallic lophi; pseudepiphallic parameres strongly bent at interno-anterior end.
Juvenile ( Fig. 3 View FIGURE 3 ). Colouration pale yellow or white, characterised by numerous dark stripes all around body in early instars; antennae with 2 wide whitish rings as in adults.
Measurements (in mm, means in brackets, N = 5 males). BL = 13.9–15.3 (14.6), PronL = 1.8–2.1 (1.9), PronW = 2.7–3.1 (2.9), FWL = 10.1–11.0 (10.4), FWW = 3.8–4.1 (3.9), HWT = 4.5–7.6 (6.6), FIIIL = 13.4–15.5 (14.5), FIIIW = 2.5–2.9 (2.6), TIIIL = 13.5–15.5 (14.4), Stridulatory teeth = 94–95.
Comparison with N. vittatus View in CoL View at ENA
Larger, with dark shiny lateral field of FWs in both sexes, below a vivid yellow band; face yellow with dark brown to black mouthparts (entirely yellow with a few dark spots in N. vittatus ); adults and juveniles ( Fig. 3 View FIGURE 3 ) are recognisable by the larger white rings on antennae (2 main rings, with 3–6 white antennomeres, while antennae are almost entirely black with in N. vittatus , with at best thin rings).
Comparison with N. musicus View in CoL View at ENA
Similar in size, but differing by dark shiny lateral field of FWs in both sexes, below a vivid yellow band; face vivid yellow (whitish in N. musicus ).
Ecology
Found in syntopy with Nisitrus vittatus , the latter species being much more abundant, and has been observed to be outnumbered by at least 10 to 1. The males and females were found inter-mixing with N. vittatus on large foliage of Leea indica along roads (forest edge) ( Fig. 4 View FIGURE 4 ). Similar to N. vittatus , this species tends to occur in open areas ( Fig. 5 View FIGURE 5 ).
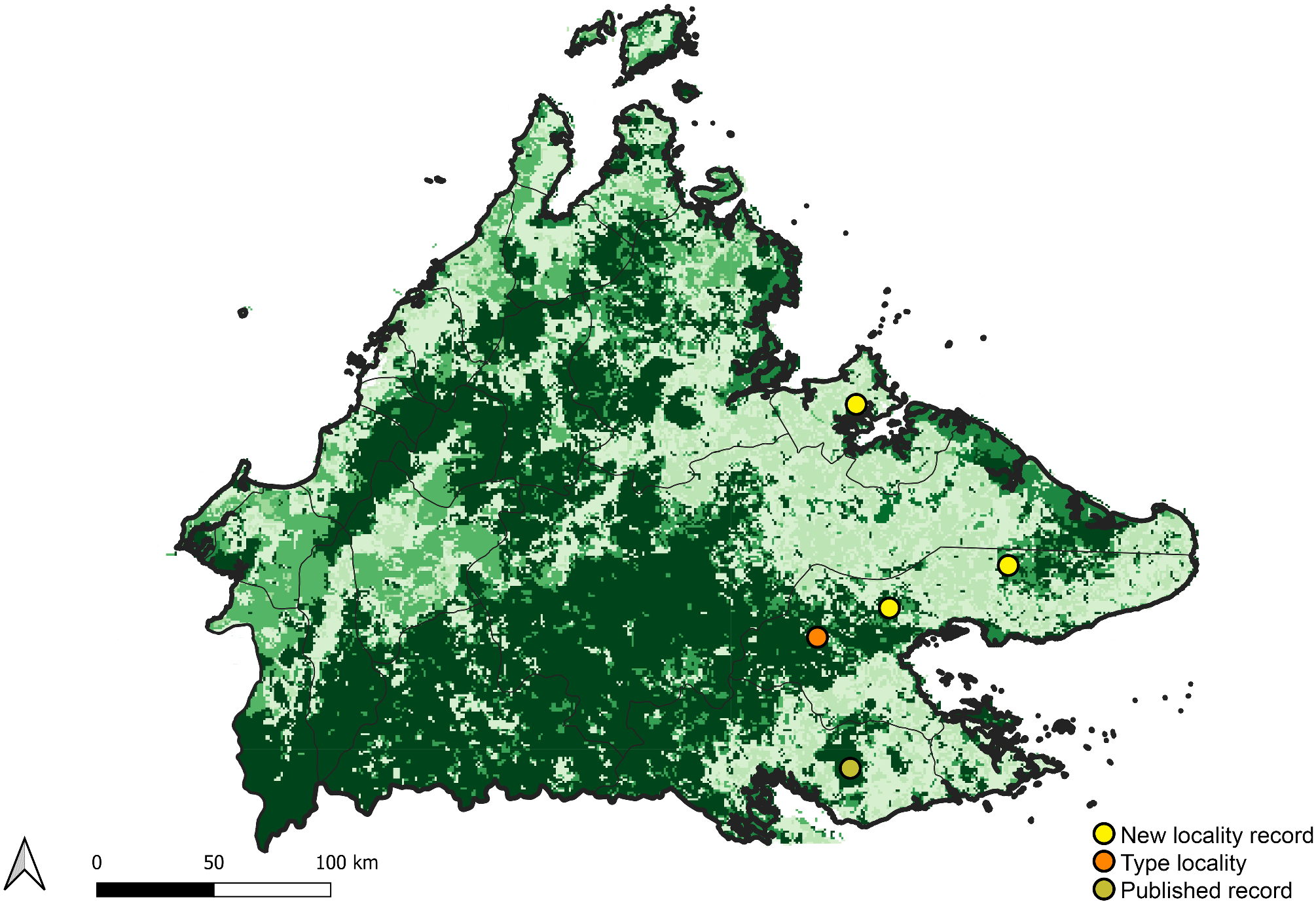
Distribution ( Fig. 6 View FIGURE 6 )
Borneo (Sabah: Danum Valley, Tawau Hill Park, Kawag Forest Reserve [new locality record], Sepilok Kabili Forest Reserve [new locality record], Tabin Wildlife Reserve [new locality record])
Calling song (9♁, 23°C, in captivity)
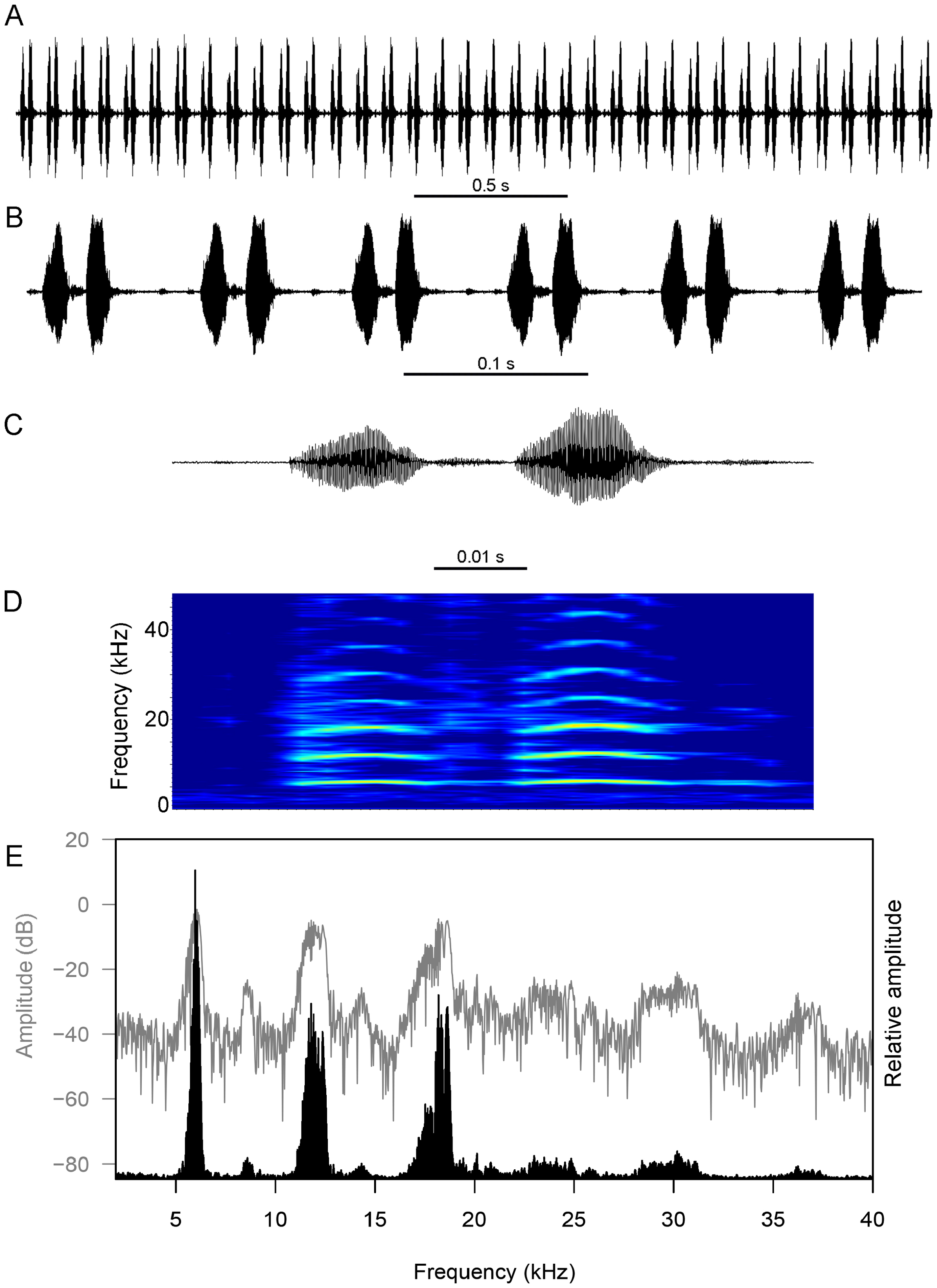
The calling song consists of long homogeneous bouts (= trill) which can last between 3 s to more than 16 min ( Fig. 7 View FIGURE 7 ). The trill is made up of a sequence of short echemes. Each echeme is made up of two syllables and has an average duration of 41.3±2.1 ms (36.6–45.9 ms). The average interval between consecutive echemes is 50.1±5.2 ms (42.6–62.1 ms). On average, the first syllable has a slightly shorter duration of 15.2±0.8 ms (13.2–17.3 ms) than the second syllable with a duration of 16.0±1.2 ms (12.6–18.7 ms). The average interval between the two syllables is 10.0±1.0 ms (7.9–12.3 ms). The call spectrum consists of a series of harmonics with three distinct peaks in the first three harmonics: first harmonic (fundamental frequency) at 6.13±0.14 kHz (5.72–6.47 kHz), which is typically also the dominant frequency; second harmonic at 12.21±0.31 kHz (11.34–12.84 kHz); third harmonic at 18.32±0.43 kHz (16.97–19.22 kHz). While second and third harmonics are not dominant, their amplitude is high and sometimes almost co-dominate with the fundamental frequency.
The calling song of N. danum is very distinct from that of the syntopic species, Nisitrus vittatus (see Tan et al., 2021a), by the echeme structure being made up of two syllables, rather than three syllables. The echeme duration is also distinctly longer in N. danum than in N. vittatus (average duration = 41.3 ms vs. 32.5 ms). Consequently, the syllable durations are also longer in N. danum than in N. vittatus . The interval between consecutive echemes is also distinctly longer in N. danum than in N. vittatus (average duration = 50.1 ms vs. 22.7 ms). Although the frequencies of N. danum is lower than that of N. vittatus , it could be an artefact of the lower ambient temperature when N. danum was recorded (23°C for N. danum vs. 27°C for N. vittatus ).
Despite morphological similarities between N. danum and N. musicus (also from Sabah, but specifically from Mount Kinabalu), there are also differences in their calling song. Both species have similar echeme structure made up of two syllables, but the echeme duration is distinctly longer in N. danum than in N. muscius (average duration = 41.3 ms vs. 29.7 ms). While the durations of the first and second syllables in each echeme are somewhat similar in N. danum (15.2 ms and 16.0 ms), those in N. musicus are drastically different (7.5 ms and 18.7 ms).
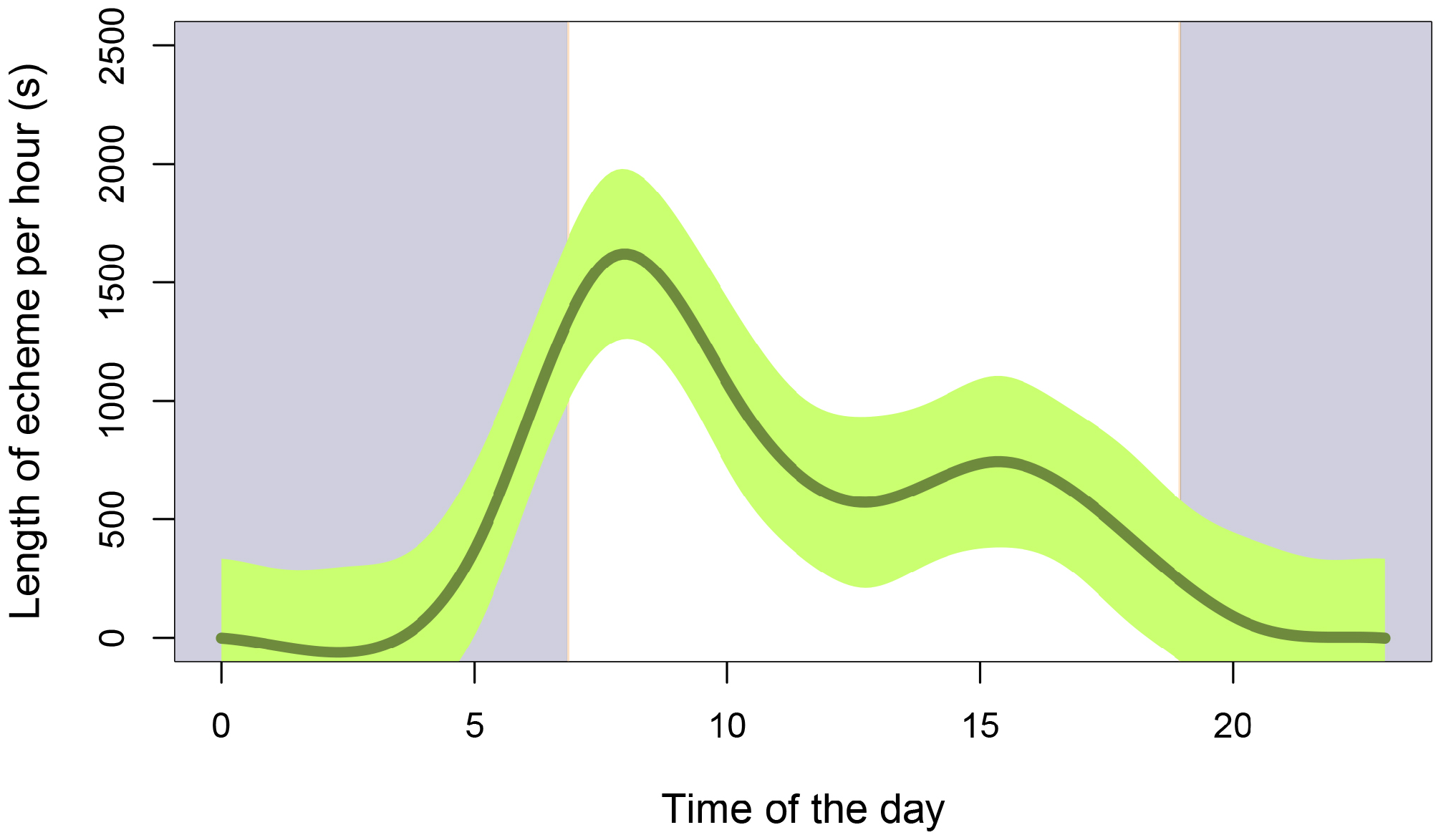
We found that N. danum is also a diurnal species, similar to other Nisitrus ( Fig. 8 View FIGURE 8 ). The circadian rhythm of the calling activity was as follows: calling began at 6:00h in the morning and its activity increases to a maximum at 8h in the morning (amount of sound produced per hour = ca. 1683 s). This follows by a drop in calling activity. A second smaller peak in calling activity (amount of sound produced per hour = ca. 915 s) was detected in the afternoon around 16:00h. Most calling activity ceases at 19:00h in the evening. The two diurnal peaks were also observed in N. vittatus (see Tan & Robillard, 2021b) but they differ in that the highest peak for N. danum was in the morning whereas that for N. vittatus was in the afternoon.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |