
Formica suecica Adlerz, 1902
|
publication ID |
https://doi.org/10.5281/zenodo.5392741 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB87B2-FFAA-F142-4F1B-FC3FFB10FDF3 |
|
treatment provided by |
Marcus |
|
scientific name |
Formica suecica Adlerz, 1902 |
| status |
|
Formica suecica Adlerz, 1902 View in CoL
TYPE LOCALITY. — Madelpad, Island of Alnön, near Sundsvall, Sweden.
MATERIAL EXAMINED. — 2 males, 1 queen, 6 workers ( NHRM), 6 queens, 6 workers ( MHN) [investigated]. GEOGRAPHIC ORIGIN OF THE MATERIAL STUDIED. — The numerically evaluated 92 specimens ( 62 workers, 28 queens, 3 males) came from Sweden 48, Finland 5, and Austria 30. Total number of specimens seen> 150.
DESCRIPTION
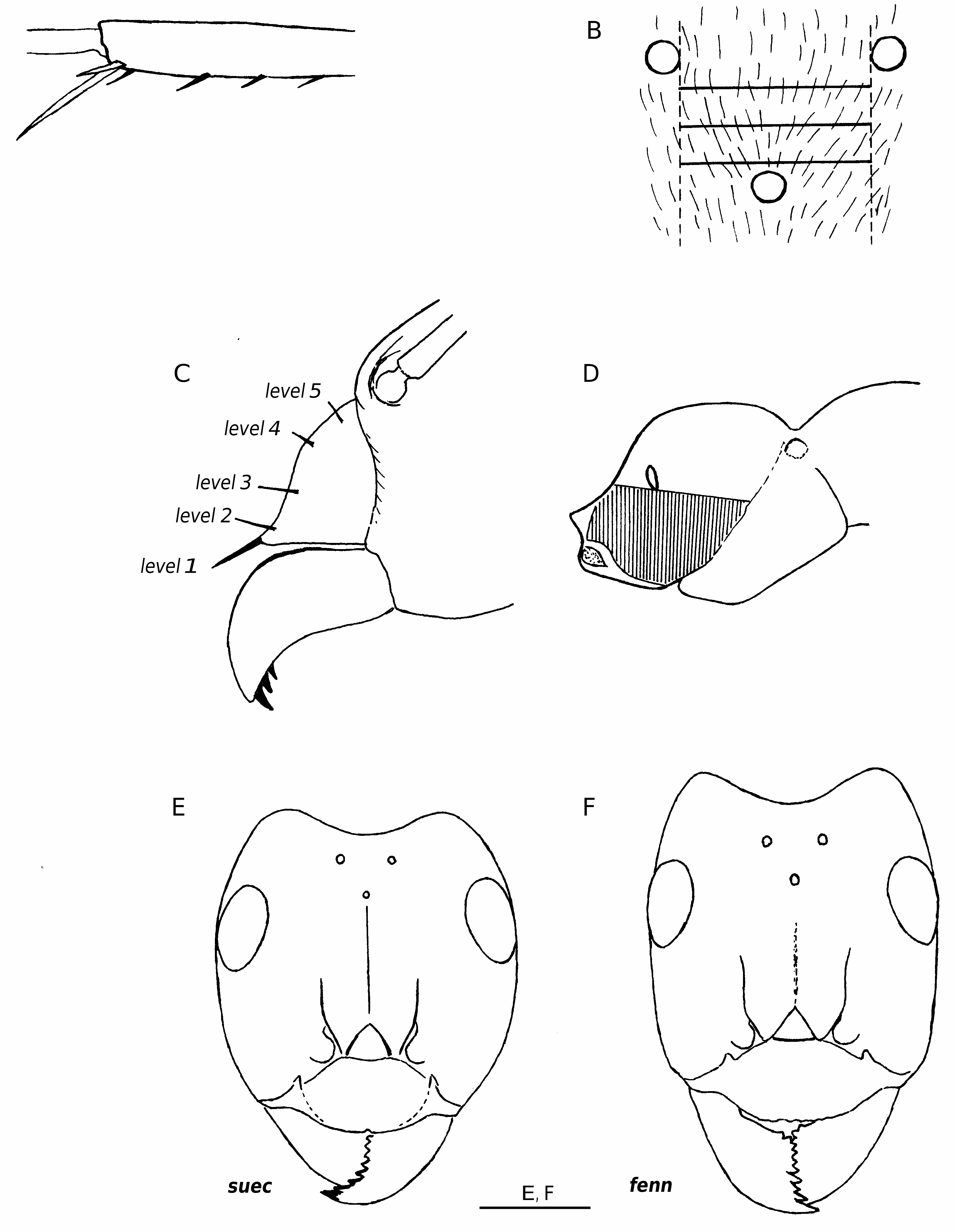
Worker ( Figs 1E View FIG ; 2 View FIG )
Relatively large (CL 1345 ± 44, 1214-1427; CW 1327 ± 55, 1177-1463). Head significantly broader, and with distinctly more convex sides and more rounded occipital corners than in other Coptoformica species ( Fig. 1E View FIG ); CL/CW 1.015 ± 0.019, 0.951-1.066). Dorsum of head, in particular area between and near the frontal carinae, mildly shining. Scape rather long (SL/CL 1.033 ± 0.020, 0.981-1.075). Setae sparsely distributed over the whole clypeus (ClySet 4.0 ± 0.0,4-4). Lateral semi-erect setae in the ocellar triangle frequently present (OceSet 72%). Eye hairs fully absent or few minute hairs present (EyeHL 6.2 ± 2.0, 0-13). Pubescence hairs in the occellar triangle short and very dense (sqrtPDF 4.38 ± 0.21, 3.96-4.86). Craniad profile of forecoxae always with semierect setae (nCOXA 4.81 ± 1.47, 1-8). Single setae on lateral metapleuron and ventrolateral propodeum may be present (nMET 1.30 ± 0.88, 0-3.0). Outer edge of the hind tibial flexor side with suberect to subdecumbent first order setae, second order setae absent ( Fig. 2 View FIG , nHTFL 6.94 ± 1.02, 4.5-9.5). Erect setae on gaster tergites always beginning on the first tergite (TERG 1.0 ± 0.0, 1-1). Pubescence on first gaster tergite very dense (sqrtPDG 5.25 ± 0.38, 4.61-5.83). Dorsal head never with a contrasting reddish and blackish pigmentation pattern as usually seen in the other species; head colour normally homogeneous reddish, more rarely dark reddish brown.
Queen
Standing setae always well-separable from pubescence that is fully appressed on whole body. In most of the size data smaller than worker (CL 1252 ± 22, 1214-1285; CW 1280 ± 24, 1232-1318; ML 2061 ± 67, 1960-2175). Head not significantly broader than in other species (CL/CW 0.978 ± 0.010, 0.951-0.994), scape long (SL/CL 0.942 ± 0.014, 0.905-0.966). Setae sparsely distributed over the whole clypeus. Erect setae in the ocellar triangle usually present. Eye hairs absent or very minute (EyeHL 6.9 ± 2.6, 0-14). Pubescence in the occellar triangle short and dense (sqrtPDF 3.98 ± 0.29, 3.58-5.03). Occipital corners of head with appressed pubescence (OccHD 0.6 ± 3.1, 0-16). Dorsal head surface shining (GLANZ 2.39 ± 0.35, 2.0-3.0). Craniad profile of forecoxae always with semierect setae (nCOXA 7.93 ± 1.32, 5.0-10.0). Dorsal mesosoma always with standing setae (MnHL 98.2 ± 10.3, 82-123). Outer edge of the hind tibial flexor side with suberect to subdecumbent first order setae, second order setae absent (nHTFL 6.15 ± 0.91, 4.5-8.0). Erect setae on
)(n
gaster tergites always beginning on the first tergite (TERG 1.0 ± 0.0, 1-1). Pubescence on first gaster tergite very dense (sqrtPDG 4.75 ± 0.41, 4.24-5.67). The dark brown body colour has often a yellowish tinge.
TAXONOMIC COMMENTS AND
DIFFERENTIAL DIAGNOSIS
F. suecica is the most constant species of the subgenus ( Tables 7; 9), showing a unique character combination. The disjunct Fennoscandian and Alpine populations show no differences (Glaser & Seifert 1999). The discrimination from exsecta is mainly given by the much smaller EyeHL and CL/CW, the absence of second order setae on hind tibia flexor profile ( Fig. 2 View FIG ), the smaller sqrtPDF, the fully appressed pubescence on the occipital corners, and the more shining head surface. F. exsecta workers may sometimes approach suecica in a single character: either the head length index may be reduced, the head colour can vary to homogeneous reddish, or eye hairs may be reduced. However, among> 1000 exsecta workers seen, not a single specimen showed a character combination inducing confusion with suecica .
BIOLOGY AND DISTRIBUTION
Geographic range
Formica suecica represents a boreo-alpine species. The known European distribution includes entire Fennoscandia between 56°N and 68°N, Esthonia, and the region of Arkhangelsk. The only known Siberian site at Tevris ( 57.31°N, 72.20°E) seems disjunct but there is no argument that suecica is not continuously distributed from N Europe to Siberia within the zone of boreal coniferous forests. It is rarely collected and the insufficient knowledge of its character differences to exsecta could have led to under-
A
C
recording. F. suecica has been considered as exclusively boreal ant, but recently a disjunct Alpine population was discovered in the upper Ötztal/ Austria at 1940-2200 m (Glaser & Seifert 1999).
Habitat selection
In Fennoscandia, suecica was frequently found at the sunny margins of bogs in the transition zone from turf to mineralic soil (42 nests), less frequently in sun-exposed heath-like habitat patches within woodland stands of Picea abies , Abies alba , or Betula on mineralic soil (23 nests), and exceptionally in a meadow (one nest) (data of Forsslund 1949 and Agosti 1989 combined with my own results). The only known alpine population at Obergurgel in the Ötztal/ Tirol was found in habitats with very cold local climate. One habitat, a 19°N-exposed alpine heath with Empetrum nigrum , Vaccinium myrtillus , Rhododendron ferrugineum, Juniperus sibiricus, and scattered old trees of Pinus cembra , has a mean annual air temperature of 2.1 °C with snow melting usually in late May. The other main habitat was N- or W-exposed alpine heath of Vaccinium myrtillus , Rhododendron ferrugineum , Vaccinium vitis-idaea , and grasses.
Status as threatened species
The population in the upper Ötztal is obviously very strong. Suecica is here the only Coptoformica species and codominant with Formica lugubris and paralugubris . In the Red List of Austrian ants it should have the status R (isolated population that is not directly endangered).
Colony foundation
Evidence as to which host species is used by the very small suecica queen for socially parasitic nest foundation is very scant. However, Formica lemani must be one of the hosts since it was the only Serviformica found in the Ötztal sites. Furthermore, lemani is the most abundant Serviformica in the Fennoscandian sites. Older Scandinavian reports naming “ F. fusca ” should be checked for species identity while reports on F. transkaucasica as occasional host seem credible. Colonies are mainly monodomous (> 90%). The tendency to distribute by nest splitting and to build up polycalic colonies is obviously very low. In three cases nests situated 1-2 m apart did not behave aggressively after experimental mixing of their workers, which suggests occasional polygyny/polycaly.
Nest construction
Regular spheric mounds of suecica preferentially constructed with finely cut pieces of grasses as typical for Coptoformica were observed in only 5% of the 66 Fennoscandian nests but were not found in any of the 32 nests in the Ötztal. The nest is usually very similar to that of Formica truncorum or sanguinea; it is often an irregular heap of litter around a tree stump or dead log in which galleries and nest chambers are excavated by the ants. The retrieved materials are mainly leaves of Ericaceae , conifer needles, or bark pieces. Parts of the mound may consist of finely-cut grass pieces. Sometimes litter-hypolithion nests are observed. In the cold, N-exposed sites of the Ötztal very special nest constructions were found. Several nests were equipped with flat solar collectors made of dark litter particles. One nest was a combination of a disc-shaped solar collector of 100 cm diameter and a 200 cm long extension of litter along a rock crevice. Another nest was situated in a huge Pinus cembra stump of 180 cm diameter in the centre of which a disc of 80 cm diameter was built with dark organic material. A third nest type in the Ötztal was a big Vaccinium bult with the inner 40 cm of diameter being a cylinder of humous material that was roofed by a moss-litter layer.
A
C B
Development and microclimatic requirements
The population in the upper Ötztal seems to have a delayed phenology with quick brood development. During a visit on 22 June 1997, the majority of nests did not contain any broods; few nests contained eggs and very few nests some eggs plus small larvae. The syntopic Formica lemani nests simultaneously had always eggs, often larvae, and sometimes even pupae. On 9 and 10 August 1997, six nests of suecica containing alates were dug out, some of which had just eclosed from the pupae. The developmental time from the egg to the alate is thus estimated as only 50-55 days in this cold alpine environment which is equal or shorter than that of Formica bruni alates under the conditions of xerothermous habitats in the Swiss Valais. The rapid brood development is most probably an adaption to the short vegetation period in the Ötztal. Provided the freshly eclosed alates need not much feeding and harden their cuticule within few days, the nuptial flights could begin in mid August. Nevertheless this population suffers a high risk of completely missing the nuptial flight after cold summers or due to earlier beginning of winter.
Demography of nests and colonies
No concrete information is available. The nest populations are normally rather small but the largest nests seen should have contained> 50000 workers.
Swarming
The flight occurs in S Sweden in mid or late July, in N Sweden in mid or late August, and is expected to occur in the upper Ötztal not before mid August. Modalities and timing of swarming are unknown.
Intra- and interspecific behaviour
At least the workers of smaller suecica nests are not aggressive during disturbance by the collector and tend to escape. Chorology in Fennoscandia and the Alps suggests inferiority of suecica in competition with exsecta in the majority of habitats on mineralic soil and that suecica is displaced to marginal habitat patches. In montane and subalpine habitats of Graubünden and Tirol, exsecta is the absolutely dominating Coptoformica species. The persistence of one isolated and populous suecica population in the Ötztal might possibly be explained by a better adaptation of brood development to the very cold local climate and short vegetation period.
TAXA WITH INSUFFICIENT DESCRIPTIONS AND UNAVAILABILITY OF TYPES
The existence or location of the types in the following W Palaearctic taxa is unknown and their descriptions are lacking diagnostic information. As a consequence, these taxa are incertae sedis and students of Coptoformica should not speculatively use these names in any context unless types are discovered and reliably identified.
Formica exsecta var. exsectorubens Ruzsky 1905
TYPE LOCALITY. — Siberia.
Formica exsecta var. wheeleri Krausse 1924
TYPE LOCALITY. — Eberswalde, Germany.
KEY AND COMPARATIVE TABLES TO WORKERS
For a successfull use of setae characters, pay attention to remains or insertion points of detached setae. The dichotomuous key is restricted to Europe. For Asiatic species use Tables 6 and 7.
1a. Eyes usually with conspicuous hairs, sometimes reduced in a big portion of specimens within a nest sample, but nest means of EyeHL always> 8 µm .................... 2
1b. Eyes without or only with microscopically short hairs, nest means of EyeHL always <8 µm ................................................................................................................ 4
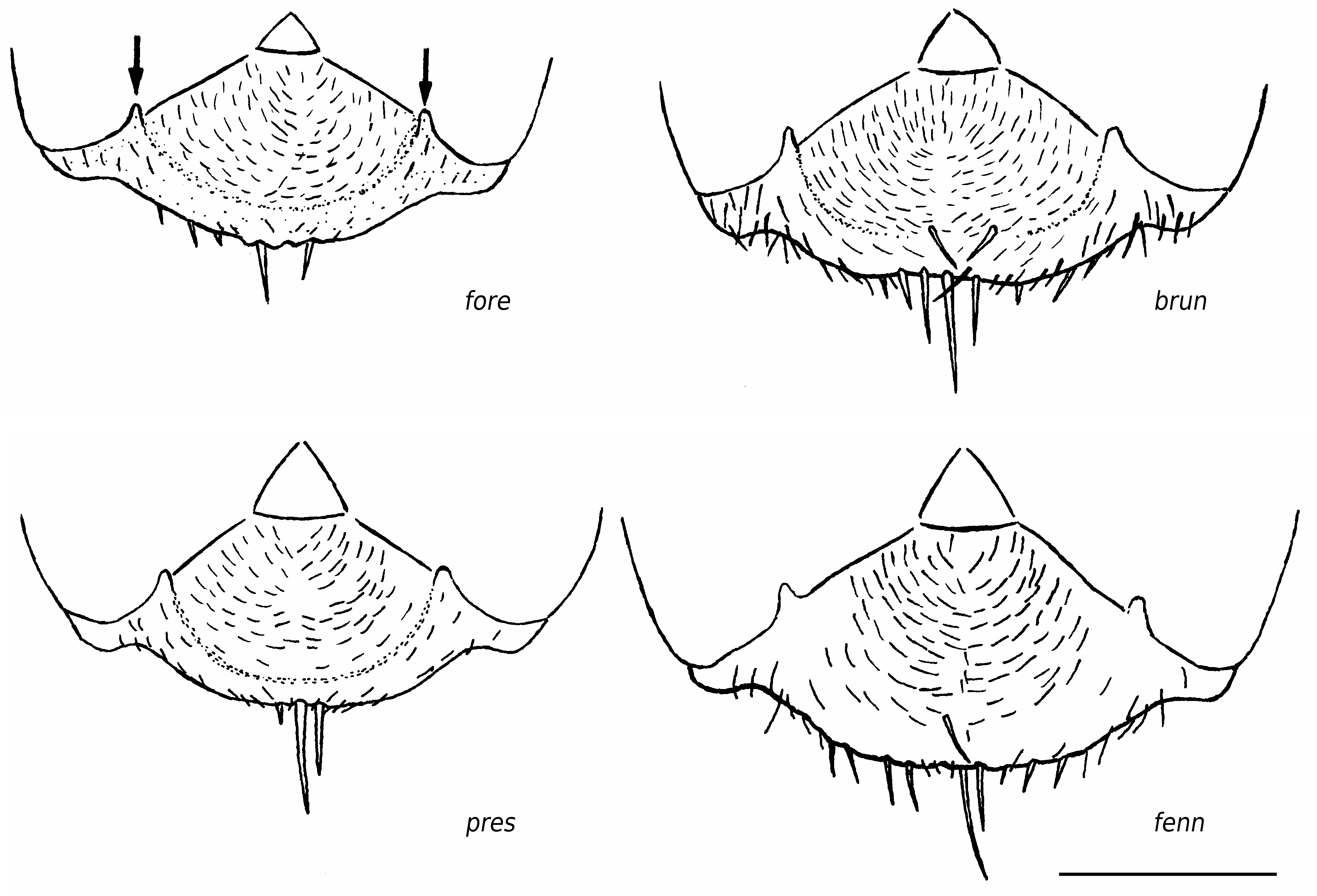
2a. Standing setae usually beginning on the first or second gaster tergite, nest means of TERG 1.0-2.3. Scattered setae on clypeus usually distributed from anterior margin to center at least; nest means of ClySet 2.5-5. Craniad profile of forecoxae usually with numerous semierect setae; nest means of nCOXA 2.5-18 ...................... exsecta View in CoL
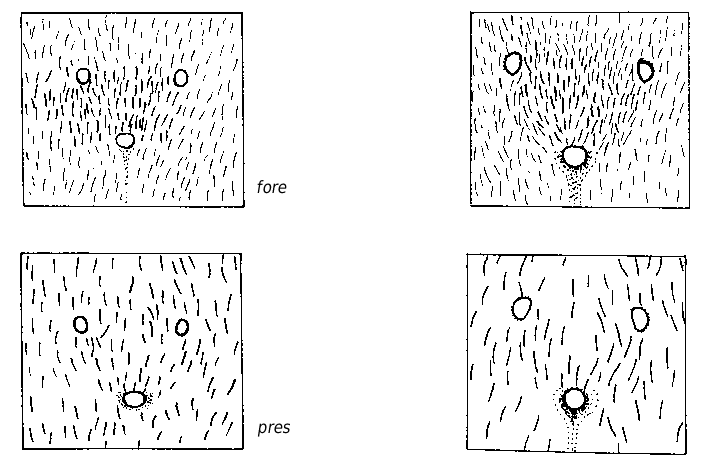
2b. Standing setae usually beginning on the third or fourth gaster tergite; nest means of TERG 2.3-4.5. Setae on clypeus usually restricted to anterior margin and single second level setae ( Fig. 3 View FIG : brun; fenn); nest means of Clyset 1.0-2.0. Craniad profile of forecoxae without or only few semierect setae; nest means of nCOXA 0-1.5 .... 3 3a. Pubescence in the ocellar region very dense ( Fig. 4 View FIG ); nest means of sqrtPDF <5.0. Pubescence on dorsum of first gaster tergite very dense; nest means of sqrtPDG <6.9. Setae on outer edge of hind tibial flexor profile rather few and usually resticted to distal half ( Fig. 2 View FIG : brun); nest means of nHTFL <5.5. Head not elongated and occipital corners less acute, nest means of CL/CW 1.033 - 1.065...................................................................................................... bruni View in CoL
3b. Pubescence in the ocellar region very thin ( Fig. 4 View FIG ); nest means of sqrtPDF> 5.7. Pubescence on dorsum of first gaster tergite very thin; nest means of sqrtPDG> 6.9. Setae on outer edge of hind tibial flexor profile more numerous and usually distributed from apex to proximal third at least ( Fig. 2 View FIG : fenn); nest means of nHTFL> 5.7. Head usually elongated and occipital corners more acute ( Fig. 1F View FIG ), nest means of CL/CW 1.048 -1.085.............................................................................. fennica

| NHRM |
Naturhistoriska Rijkmuseet |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |