
Nymphaea
|
publication ID |
https://doi.org/10.1016/j.phytochem.2014.04.007 |
|
DOI |
https://doi.org/10.5281/zenodo.10561355 |
|
persistent identifier |
https://treatment.plazi.org/id/03BA87CE-FFBF-101E-FFDB-F5ED7FAEFADB |
|
treatment provided by |
Felipe |
|
scientific name |
Nymphaea |
| status |
|
2.2. Floral scent variations within Nymphaea View in CoL and Victoria
The NMDS representations of the floral scents of the selected species of Nymphaea and Victoria yielded fairly good representations of differences between individuals when two axes were maintained (stress values: S = 0.08 when considering all VOCs individually; S = 0.06 when considering VOCs pooled into chemical subclasses as proposed by Knudsen et al. (2006).
In these two ‘‘scent space’’ representations, specimens belonging to any certain species came out grouped together, with the exception of N. rudgeana that exhibited fluctuations of the relative concentrations of the two main compounds in its floral bouquet: methyl hexanoate (3) and (methoxymethyl)benzene (5) ( Fig. 4 View Fig ).
When investigating the multivariate ordination restricted to chemical subclasses, N. gardneriana and N. rudgeana came out close to the two species of Victoria , with which they share aliphatic esters and/or C5-branched chain esters, whereas the five other sampled taxa belonging to Nymphaea subg. Hydrocallis grouped together due to characteristically high emissions of aromatic esters ( Figs. 3 View Fig , 4A View Fig ). The ordination graph based on individual VOCs further divided these same five species into two separate groups in which either (methoxymethyl)benzene (5) ( N. lasiophylla , N. lingulata ) or anisole (6) ( N. amazonum subsp. amazonum , N. amazonum subsp. pedersenii and N. tenerinervia ) were dominant constituents ( Fig. 3 View Fig , 4B View Fig ).
The two species belonging to Nymphaea subg. Brachyceras ( N. caerulea and N. stellata ) grouped together, but were isolated from the other taxa on both ‘‘scent space’’ representations ( Fig. 4 View Fig ). Their floral scents are characterized by high amounts of aromatic alcohols and aromatic esters, notably benzyl alcohol (7) and benzyl acetate (8) ( Fig. 1 View Fig ). On the other hand, N. lotus (subg. Lotos ) came out in an intermediate position between species from the subgs. Brachyceras and Hydrocallis when considering the chemical subclasses of compounds in its floral scent, which is dominated by alkanes, C5-branched chain esters and aromatic ethers ( Fig. 4A View Fig ). It also came out apart in the graph based on individual VOCs due to a more specific scent composition, notably by the presences of pentadecane (9) (shared with species belonging to subg. Brachyceras) and 2,5-dimethoxytoluene (10) ( Fig. 4B View Fig ).
3. Discussion
The floral scents of the investigated species of Nymphaea subg. Hydrocallis were characterized by remarkably simple blends. Not only were they comprised by few constituents (12 or less in all analyzed samples), but they also each contained only one or two prominent compounds that alone accounted for over 95% of total scent discharge ( Fig. 3 View Fig ; Table 2 View Table 2 ).
Contrastingly, most of the plant taxa pollinated by highly specialized insect pollinators are characterized by complex floral scent blends, easily constituted by 80 or more volatile compounds ( Dobson, 2006; Knudsen et al., 2006). Often, such intricacy is needed for triggering pollinators’ behavioral responses, as several compounds might act synergistically within a very narrow concentration range in the floral scent blend in order to be attractive ( Raguso, 2008).
⁄ Main constituent in the floral scent of Nymphaea gardneriana. w Present in high relative proportion in the scent of Nymphaea gardneriana and N. lasiophylla. † Present in high relative proportion in the scents of Nymphaea amazonum amazonum; N. amazonum pedersenii, N. garneriana, N. lasiophylla, N. lingulata, N. tenerinervia, N. rudgeana. 1 Gottsberger et al. (2012). 2 Dötterl et al. (2012). 3 Maia et al. (2012). 4 Maia et al. (2013).
Maia et al. (2012) speculated that it might be strategically common among angiosperms pollinated by cyclocephaline scarabs that one to few compounds are applied as specific communication channels to attract pollinators. Cyclocephaline scarabs from genera Cyclocephala and Erioscelis are attracted to pure standards of compounds found in floral scents ( Table 3 View Table 3 ). As a matter of fact, all known attractants of pollinator cyclocephaline scarabs have been recovered as major constituents in the fragrances of flowers and/ or inflorescences of their preferred plant hosts ( Dötterl et al., 2012; Gottsberger et al., 2012; Maia et al., 2012, 2013). In such a fashion, the selectivity cues involved in these plant-pollinator associations could be ultimately linked to the recognition of unique chemical signals, warranted by the emission of rare floral scent volatiles in high concentrations. Although the emission of VOCs by flowers of Nymphaea subg. Hydrocallis was more pronounced during the pistillate phase of anthesis, flowers at the staminate phase were still highly fragrant ( Table 2 View Table 2 ). Because there is little difference in floral scent composition between phases as well (with the exception of N. rudgeana ), it is likely that attractive VOCs induce the scent-oriented pollinators to fly indiscriminately between pistillate and staminate fragrant flowers. Such behavior was observed by Gottsberger et al. (2012) when studying the reproductive ecology of Magnolia ovata ( Magnoliaceae ), a strategy that enhances pollen mixing and effective cross-pollination of the self-compatible species. The distinct relative proportions of the most abundant VOCs in the floral scent N. rudgeana ( Fig. 3 View Fig ; Table 2 View Table 2 ) require further investigation, but might be linked to differences in pollinator attractiveness during the consecutive phases of anthesis. For example, it has been shown that the flowering cones of cycads ( Cycadaceae ) emit different floral scent compositions in the course of anthesis. In the first day, floral scent is attractive to pollinators; in the second, it is repulsive, forcing the pollinators to leave the cones loaded with pollen ( Terry et al., 2007).
Aromatic ethers, the main VOCs emitted by the investigated species of Nymphaea subg. Hydrocallis , are allegedly derived from the metabolism of phenylalanine in plants ( Dudareva et al., 2013). They are exceptionally frequent in floral scents of angiosperms pollinated by cyclocephaline scarabs and have long been associated with their attraction ( Dobson, 2006; Kite et al., 1991), an assumption which has recently received solid support from studies with aroid pollinators ( Dötterl et al., 2012) ( Table 3 View Table 3 ). The floral scents of both subspecies of N. amazonum , as well as that of N. tenerinervia exhibited extremely high relative amounts of anisole (6), an aromatic ether never before recorded as a major constituent in floral scents. Its occurrence was actually restricted to seven early-diverging angiosperm families, notably Araceae , Arecaceae , Magnoliaceae and Nymphaeaceae ( Knudsen et al., 2006) . (Methoxymethyl)benzene (5), like anisole (6), is also a rare floral scent constituent and was previously only known from five angiosperm families, mostly as a trace or minor constituent ( Knudsen et al., 2006). It is a dominant constituent in the scents of N. lingulata and N. lasiophylla , as well as a key compound in the more complex blend of N. rudgeana .
When applied to baits, undiluted aliquots of the structurally similar p -vinylanisole (11) ( Fig. 2 View Fig ), the most abundant compound in the floral scent of Philodendron selloum , yielded captures of beetles of Erioscelis emarginata ( Scarabaeidae, Cyclocephalini ), specialized pollinators of this plant in Central Brazil ( Dötterl et al., 2012) ( Table 3 View Table 3 ). The fact that anisole (6) and (methoxymethyl)benzene (5) appear as absolute dominant constituents in the scents of six species of Nymphaea subg. Hydrocallis leads us to assume that they might also be involved in specific pollinator attraction.
One of the main constituents in the floral scents of N. lasiophylla and N. gardneriana , methyl 2-methylbutanoate (4), a chiral ester likely derived from the plant metabolism of L- isoleucine ( Rowan et al., 1996), was also isolated from flowers of the two species of Victoria ( Kaiser, 2006; Kite et al., 1991) ( Fig. 3 View Fig ; Appendix Table 2 View Table 2 ). Kite et al. (1991) also speculated that this compound could be implicated with the attraction of cyclocephaline scarabs, an assumption later confirmed by Gottsberger et al. (2012) in a study with male and female Cyclocephala literata , the sole pollinators of M. ovata ( Magnoliaceae ) ( Table 3 View Table 3 ).
Yet another compound previously identified as a major constituent in the floral scent of V. amazonica is methyl hexanoate (3) ( Kaiser, 2006; Kite et al., 1991) ( Fig. 3 View Fig ; Appendix Table 2 View Table 2 ), also found in high concentration in flowers of N. rudgeana and N. gardneriana during anthesis ( Fig. 3 View Fig ; Table 2 View Table 2 ). Although there is no evidence of its role in pollinator attraction, it has been isolated from males of different species of Drosophila and is assumed to be part of their aggregation pheromones ( Bartelt et al., 1986, 1988).
None of the main compound subclasses isolated in night-blooming Nymphaea subg. Hydrocallis and Victoria were recovered from day-blooming species of Nymphaea subg. Brachyceras ( Figs. 3 View Fig , 4B View Fig ; Appendix Table 2 View Table 2 ). Such clear distinction in the floral scent compositions between these groups of taxa suggest they could be related to pollinator selection, an evolutionary trend that has been clearly demonstrated in study cases involving several plant-pollinator systems ( Jürgens et al., 2000; Knudsen and Tollsten, 1993; Steenhuisen et al., 2013). A broader screening of VOCs among all genera and subgenera of the Nymphaeaceae , studied under the light of phylogenetic relationships, would yield interesting insights into the evolution of floral scents and pollination systems in the family (Lima et al., unpublished data).
4. Concluding remarks
The floral scents of Neotropical night-blooming Nymphaea belonging to the subgenus Hydrocallis were investigated for the first time in the present study. According to the most recent phylogeny of the Nymphaeaceae , it appears that nocturnal anthesis and specialization towards cyclocephaline scarab-pollination was a key evolutionary move within species of the water lily family, as it groups together not only Nymphaea subgenera Lotos and Hydrocallis , but also the genus Victoria ( Löhne et al., 2008) . Scent preference bioassays with volatile compounds isolated from the floral scents of Nymphaea subg. Hydrocallis should further clarify their specific role in the attraction of cyclocephaline scarabs.
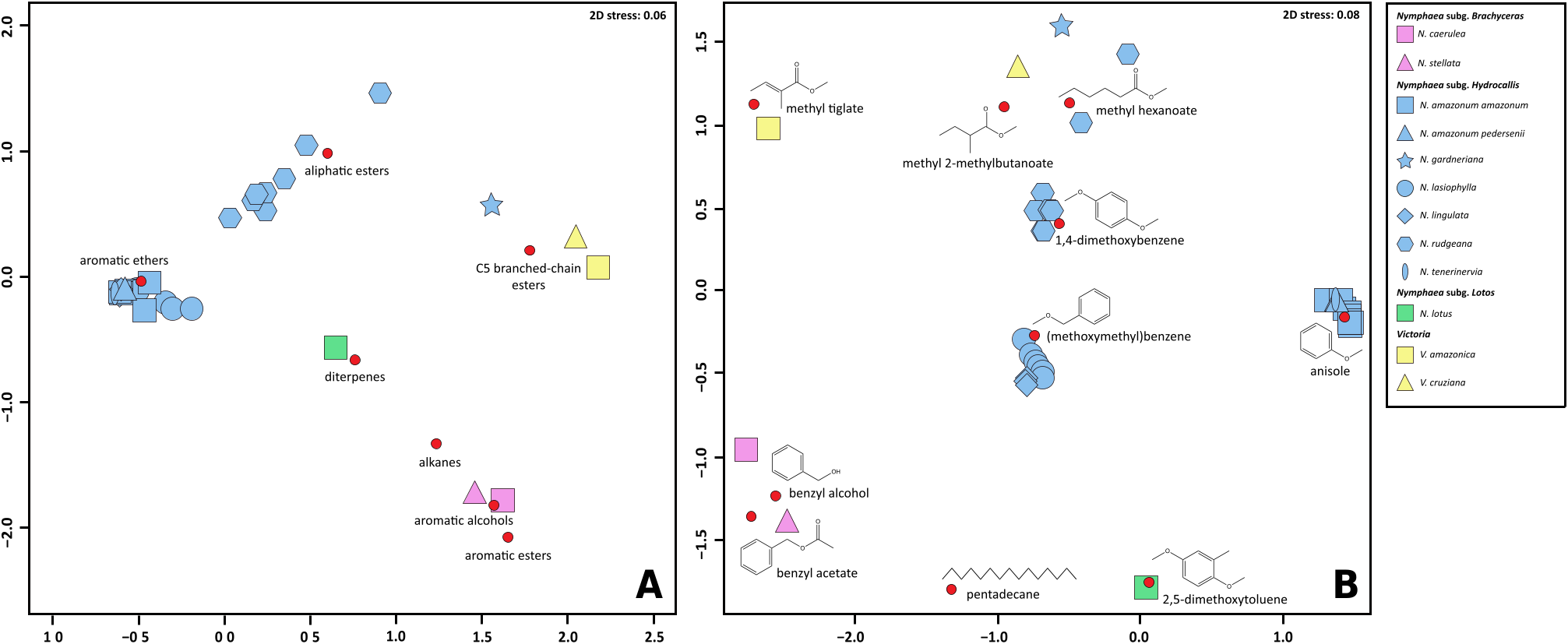
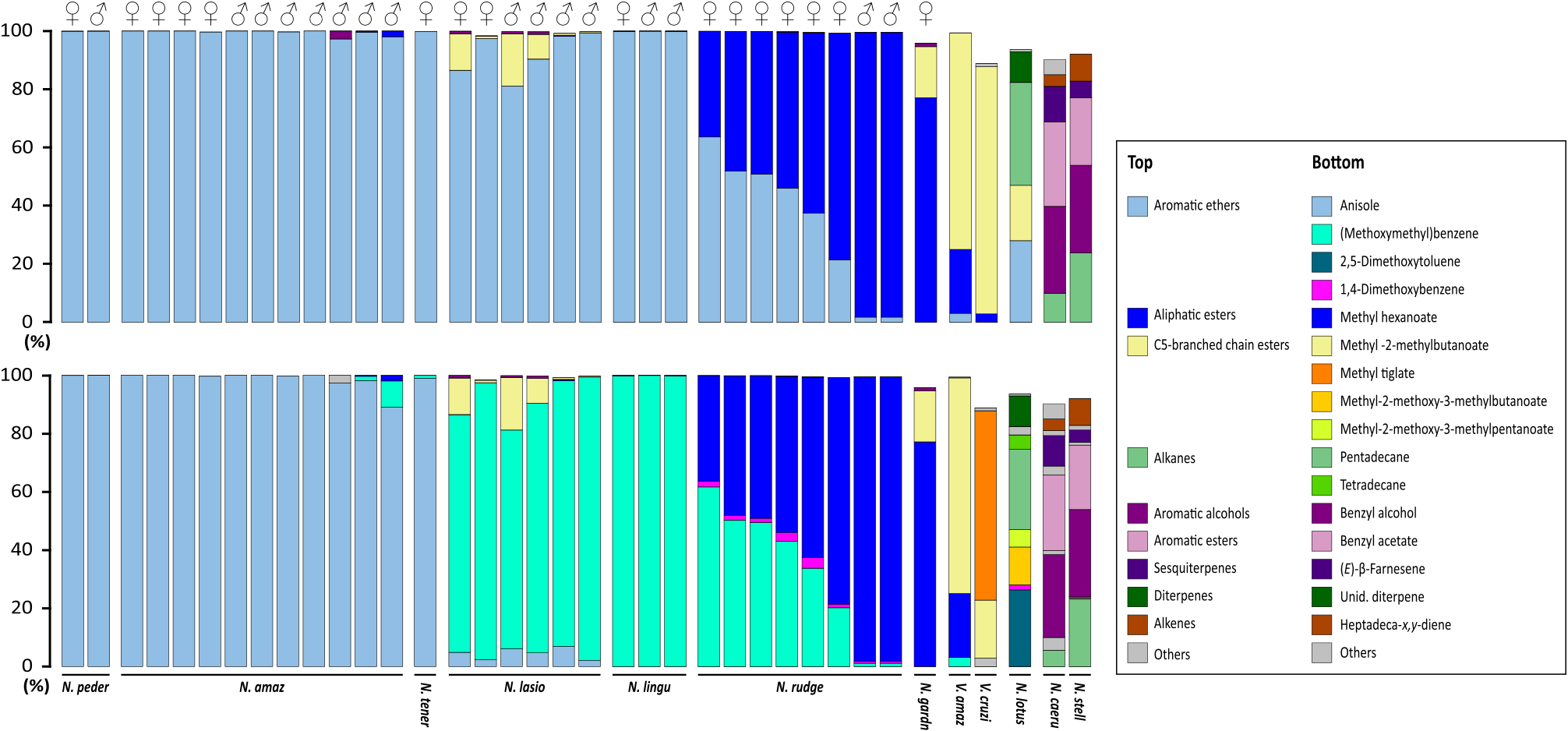

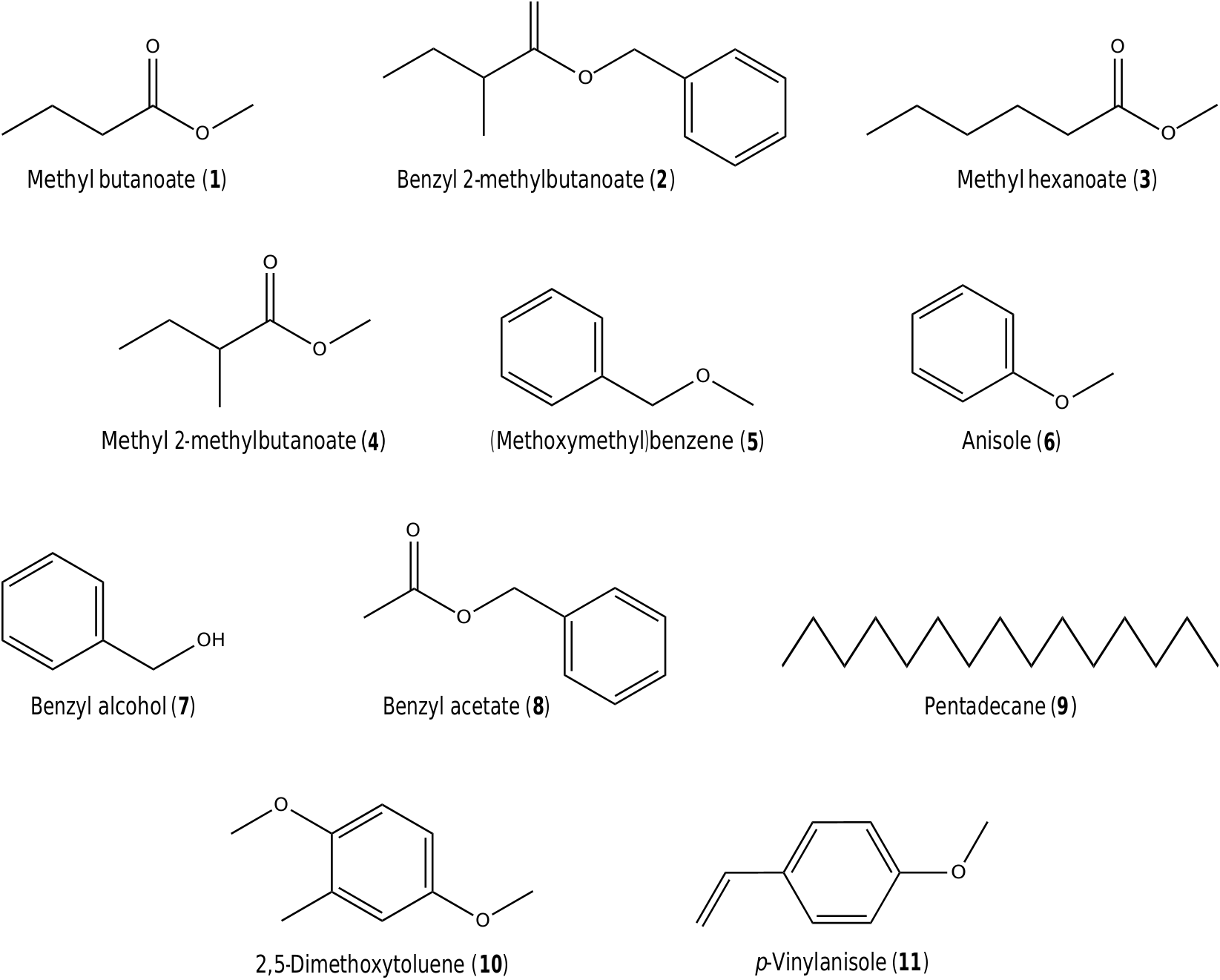
Table 3 Compounds known to elicit attractive behavior response from pollinator cyclocephaline scarab beetles (Scarabaeaidae, Cyclocephalini).
| Attractant | Compound subclass | Related plant species (relative% in scent) | Attracted pollinators |
|---|---|---|---|
| C5-branched chain ester w | Magnolia ovata (>80%) | Cyclocephala literata 1 | |
| Aromatic ether † | Philodendron selloum (>65%) | Erioscelis emarginata 2 | |
| Sulfur-containing comp. | Caladium bicolor (>35%) Annona spp. (>95%) | Cyclocephala celata 3 Cyclocephala vestita 3 | |
| Aliphatic ketone | Taccarum ulei (>70%) | Cyclocephala celata, Cyclocephala cearae 4 |
| A |
Harvard University - Arnold Arboretum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |