
Cladocarpus pennatus, Galea, 2020
|
publication ID |
https://doi.org/ 10.5852/ejt.2020.615 |
|
publication LSID |
lsid:zoobank.org:pub:637FC87F-13B5-4B32-BC52-11A9B30ECF1D |
|
DOI |
https://doi.org/10.5281/zenodo.3718317 |
|
persistent identifier |
https://treatment.plazi.org/id/7E29B1BC-C12D-43BE-8D91-0BB238C6CDBA |
|
taxon LSID |
lsid:zoobank.org:act:7E29B1BC-C12D-43BE-8D91-0BB238C6CDBA |
|
treatment provided by |
Plazi |
|
scientific name |
Cladocarpus pennatus |
| status |
sp. nov. |
Cladocarpus pennatus View in CoL sp. nov.
urn:lsid:zoobank.org:act:7E29B1BC-C12D-43BE-8D91-0BB238C6CDBA
Figs 6 View Fig C–E, 8–9; Table 2 View Table 2
Diagnosis
Cladocarpus with large, fan-shaped colonies, with strongly fascicled stems and side branches. Nodes rather indistinct, internodes with a proximal, frontal nematotheca, a cladial apophysis with its coneshaped nematotheca, as well as a fronto-axillar nematotheca. Cladia close to one another, divided into short internodes; first internode short, with a nematotheca, but no hydrotheca; remainder of cladium with normal cormidia. Hydrothecae facing backwards with respect to the antero-posterior plane of the colony; sigmoid, variously bulging abaxially from their lower halves; mesial nematotheca conspicuous, half adnate, projecting obliquely forward and upward (shorter and strongly pointing laterally on first cormidium); hydrothecal aperture with a median, triangular cusp and 1–2 pairs of laterals, decreasing in size adaxially; an internal, adaxial septum plunging down into the hydrothecal lumen for half its width. Gonosome composed of gonothecae arising from the cladial apophyses and phylactocarps borne on the first cormidium of a cladium that has lost its proximalmost, nematothecate internode. Phylactocarps elongated, set with consecutive pairs of lateral nematothecae, an additional, median, distal row occasionally present. Gonothecae dimorphic, male tubular, with recurved distal end bearing large aperture flanked by two lobes, female comparatively smaller and fusiform.
Etymology
From the Latin ‘ pennātus , -a, -um ’, meaning ‘bearing plumes’, with reference to the plumose appearance of its colonies.
Material examined
Holotype
PACIFIC OCEAN • 1 colony, ca 6 × 6 cm, hydrorhiza missing, bearing male gonothecae; off New Caledonia, stn DW4720 ; 22°50ʹ S, 167°11ʹ E; 374–400 m; 19 Aug. 2016; KANACONO leg.; MNHN-IK-2015-534. GoogleMaps
Paratypes
PACIFIC OCEAN • 1 colony, ca 4.5 × 6.5 cm, hydrorhiza missing, bearing female gonothecae with welldiscerned oocytes; off New Caledonia, stn DW4720 ; 22°50ʹ S, 167°11ʹ E; 374–400 m; 19 Aug. 2016; KANACONO leg.; MNHN-IK-2015-535 GoogleMaps • two colonies, ca 8 × 4.5 cm and 5 × 4.5 cm, both without hydrorhizae, but bearing female gonothecae; off New Caledonia, stn DW4707 ; 22°46ʹ S, 167°23ʹ E; 291– 259 m; 17 Aug. 2016; KANACONO leg.; MNHN-IK-2015-537 GoogleMaps .
Additional material
PACIFIC OCEAN • 1 colony, ca 11 × 8.5 cm, bearing female gonothecae; off New Caledonia, stn CP4704 ; 22°46ʹ S, 167°19ʹ E; 365– 356 m; 17 Aug. 2016; KANACONO leg.; MNHN-IK-2015-531 GoogleMaps • 1 young, unbranched, sterile colony, 7 cm high, hydrorhiza missing; off New Caledonia, stn DW4762; 23°16ʹ S, 168°06ʹ E; 810– 805 m; 26 Aug. 2016; KANACONO leg.; MNHN-IK-2015-532 GoogleMaps • 1 colony fragment, ca 3.8 × 2 cm, bearing female gonothecae; off New Caledonia, stn DW4713; 22°47ʹ S, 167°47ʹ E; 356–380 m; 18 Aug. 2016; KANACONO leg.; MNHN-IK-2015-533 GoogleMaps • 3 colonies, ca 10 × 7 cm, 8 × 5.5 cm and 10 × 7 cm, with female gonothecae, 1 colony, 12 × 4 cm, without gonothecae; off New Caledonia, stn CP4675; 22°50ʹ S, 167°30ʹ E; 350–366 m; 13 Aug. 2016; KANACONO leg.; MNHN- IK-2015-536 GoogleMaps • 3 colonies with or without hydrorhizae and no gonothecae, ca 7.5 × 4.5 cm, 5.5 × 2 cm, and 5.5 × 7.5 cm, respectively, 1 colony, ca 5 × 6.5, without hydrorhiza and starting to reproduce (only phylactocarps but no gonothecae are present), 1 male colony, 5 × 3 cm, hydrorhiza missing, upper part of 1 large colony, ca 4.2 × 6 cm, with male gonothecae; off New Caledonia, stn DW4717; 22°44ʹ S, 167°11ʹ E; 336–361 m; 19 Aug. 2016; KANACONO leg.; MNHN-IK-2015-538 GoogleMaps • 1 long fragment, 2.7 cm, without gonothecae, with strongly bent hydrothecae and hypertrophied nematothecae; off New Caledonia, stn CP4676; 22°51ʹS, 167°30ʹ E; 383 m; 13 Aug. 2016; KANACONO leg.; MNHN- IK-2015-539 GoogleMaps • 2 colony fragments, ca 4 × 2 cm and 2.4 cm, respectively, both with male gonothecae, 1 unbranched colony, 6 cm high, without gonothecae; off New Caledonia, stn DW4679; 22°49ʹ S, 167°33ʹ E; 245–249 m; 13 Aug. 2016; KANACONO leg.; MNHN-IK-2015-540 GoogleMaps • 1 colony, ca 7 × 5 cm, with female gonothecae, 1 colony, ca 6.5 × 4.5 cm, hydrorhiza and gonothecae missing; off New Caledonia, stn DW4706; 22°47ʹ S, 167°24ʹ E; 355– 343 m; 17 Aug. 2016; KANACONO leg.; MNHN- IK-2015-541 GoogleMaps • 1 colony, ca 8 × 4 cm, without hydrorhiza, bearing male gonothecae; off New Caledonia, stn DW4708; 22°44ʹ S, 167°21ʹ E; 71– 68 m; 17 Aug. 2016; KANACONO leg.; MNHN-IK-2015-542 GoogleMaps • 1 unbranched colony, ca 5 cm high, without hydrorhiza and gonothecae; off New Caledonia, stn DW4745; 22°57ʹ S, 167°39ʹ E; 310–403 m; 23 Aug. 2016; KANACONO leg.; MNHN-IK- 2014-2594 GoogleMaps .
Description
Colonies erect, fan-shaped, up to 11 cm high and 8.5 cm wide, arising from rhizoid stolon composed of numerous branching, anastomosing, contiguous fibers firmly anchoring the whole structure on its substrate. Stems strongly fascicled, branched irregularly with a tendency to alternate, main stem supported dorsally by beam of auxiliary tubes, each provided with a longitudinal row of deeply-immersed, saccate nematothecae, having only their distal parts, bearing the apertures, protruding from the perisarc. There are no proximal oblique nodes passing through the fascicled stem; main tube with indistinct division into internodes, except for the distalmost parts of a colony, where the perisarc is thinner, and where weak transverse node delimit very short internodes; the latter composed of a proximal, frontal nematotheca, a laterally-directed cladial apophysis with its reduced conical nematotheca (with small, rounded aperture), and a second, frontal (axillar) nematotheca rather lateral to the apophysis; stem nematothecae elongated, with gutter-shaped apertures, occasionally inverted-triangular, with wide distal apertures and backwardly-rolled abaxial walls. Apophyses alternate along the stem, well-developed, delimited from the corresponding cladium by a deeply-cut, very oblique node; cladia close to one another, up to 7 mm long, slightly convex, proximally pointing upwards and distally downwards, the two rows coplanar; first cladial internode very short, ahydrothecate, but provided with a frontal, gutter-shaped nematotheca, ending in oblique node, separating it from the remainder of cladium, the latter comprising a regular succession of short internodes delimited by means of transverse nodes (100–115 µm wide); each internode 350–365 µm long, with a hydrotheca and its 3 associated nematothecae: one mesial and a pair of laterals; through a torsion of the cladium at its insertion on the corresponding apophysis, all hydrothecae face backwards with respect to the antero-posterior plane of the colony. Hydrothecae 260– 270 µm deep, S-shaped, with thick abaxial wall, with an internal septum given off from the lower ¼ of the cavity and projecting slightly downwards so as to reach about the middle of the thecal lumen, ending distally in a perisarc plug; aperture ca 160 µm wide, rim with a central, prominent, triangular, abaxial cusp, continued laterally with 1–2 additional cusps decreasing in size adaxially. Mesial nematotheca adnate for ¾ to ½ of its length to the hydrothecal base, free part 115–125 µm long, aperture adaxial and gutter-shaped over entire length of free part; lateral nematothecae long (ca 145 µm), tubular, pointing out- and upwards, adaxial wall deeply scooped out. First cladial hydrotheca with comparatively shorter mesial nematotheca, given off from below its base, and distinctly pointing towards the dorsal side of the colony. Gonosome composed of phylactocarps, each associated with one gonotheca; gonothecae given off from the cladial apophyses just above the conical nematotheca and lateral to the distalmost internodal stem nematotheca; phylactocarps given off from cladia modified proximally as follows: the proximalmost ahydrothecate internode carrying the single, frontal nematotheca is replaced by a complete cormidium delimited by very oblique nodes; its mesial nematotheca is comparatively shorter than the subsequent ones, is given off from below the hydrothecal base, and is distinctly shifted on to the dorsal side of the colony; the second cormidium, also complete, is delimited proximally by an oblique node and distally by the first transverse node of the cladium; its mesial nematotheca is more elongated than that of the first cormidium and surpasses, for a certain distance, the hydrothecal base, but is still slightly shifted laterally; the third and subsequent cormidia are similar to those of a normal hydrocladium, with well-defined, straight mesial nematothecae. The phylactocarps are given off from the first cormidium, lateral to and above the mesial nematotheca, towards the front of the colony, by means of a short, conical apophysis delimited distally by transverse node, and having a long, guttershaped nematotheca at its base; phylactocarp composed of a double row of nematothecae, 6–10 in female colonies and up to 30 in males; in the latter, the double row is often supplemented distally by a number of additional nematothecae situated dorsally in a median row. Colonies dioecious; the gonothecae occur in groups towards the distal parts of the colonies, and are given off simultaneously from numerous consecutive apophyses of the branches; male gonothecae large (ca 1395 × 460 µm), tapering below, distally narrowing to a downwardly-curved neck, with broad aperture flanked by a pair of lobes; female comparatively smaller (ca 955 × 330 µm), fusiform, with minute (ca 35 µm wide), apical aperture, despite the much larger size of the oocytes.
Remarks
Several Cladocarpus -like hydroids possess S-shaped hydrothecae provided with an adaxial septum, similar to those of C. pennatus sp. nov. However, there are morphological features enabling their specific separation, and these are summarized in Table 2 View Table 2 .
The male gonothecae of the new species recall those of Wanglaophenia longicarpa Vervoort & Watson, 2003 (see Vervoort & Watson: fig. 81h), but the carinate hydrothecae and the forked phylactocarps bearing hydrothecae ( Vervoort & Watson 2003) of the latter readily distinguishes these hydroids.
Distribution
Known only from off New Caledonia (present study).
Genus Gymnangium Hincks, 1874
Gymnangium expansum ( Jäderholm, 1903)
Fig. 10 View Fig
Gymnangium expansum View in CoL – Vervoort 1966: 165, figs 65–66. — Rees & Vervoort 1987: 163, fig. 38a–b. —
Vervoort & Watson 2003: 289, figs 68g, 69a.
Material examined
PACIFIC OCEAN • 1 colony, ca 10 cm high, lightly fascicled, fertile; off New Caledonia, stn DW5007 ; 22°12ʹ S, 159°02ʹ E; 290–750 m; 19 Sep. 2017; KANADEEP leg.; MNHN-IK-2015-515 GoogleMaps • 1 colony, ca 10.5 cm high, lightly fascicled, sterile; off New Caledonia, stn DW4777; 23°03ʹ S, 168°16ʹ E; 330–353 m; 28 Aug. 2016; KANACONO leg.; MNHN-IK-2015-516 GoogleMaps • two sterile stems, the first ca 16 cm high and monosiphonic, the second ca 20 cm high and lightly fascicled; off New Caledonia, stn DW4711; 22°47ʹ S, 167°24ʹ E; 335–338 m; 18 Aug. 2016; KANACONO leg.; MNHN-IK-2015-517 GoogleMaps • 1 colony, ca 16 cm high, lightly fascicled, sterile; off New Caledonia, stn CP4674; 22°48ʹ S, 167°29ʹ E; 311– 302 m; 13 Aug. 2016; KANACONO leg.; MNHN-IK-2015-518 GoogleMaps • 1 colony, ca 12 cm high, lightly fascicled, sterile; off New Caledonia, stn DW4672; 22°47ʹ S, 167°26ʹ E; 310– 290 m; 13 Aug. 2016; KANACONO leg.; MNHN-IK-2015-521 GoogleMaps .
Remarks
For a description of this species, refer to Vervoort (1966) and Rees & Vervoort (1987); a quite recent synonymy is given in Vervoort & Watson (2003). In these accounts, only slightly bent hydrothecae, whose nematothecae have single apical apertures, were documented, not reaching the extreme degree of curvature (aperture parallel to long axis of the internode) met with in material MNHN-IK-2015-516, in which, additionally, the mesial nematothecae display 4–6 frontal apertures, while the pairs of laterals have bifid apertures. In contrast, the material MNHN-IK-2015-515 has less curved hydrothecae (their apertures form an angle of 45° with the long axis of the internode), but their mesial nematothecae still display 2–3 frontal apertures, while the laterals possess only one. An intermediate situation is met with in sample MNHN-IK-2015-518, in which the hydrothecal apertures form an angle of ca 70° with the internode and the mesial nematothecae possess 1–2 frontal apertures, while the laterals have only one. Bifid mesial nematothecae were observed so far in specimens from Japan studied by Schuchert (2015).
In all samples examined the branching pattern is ‘trifid’, with either mono- (the 16 cm-high colony from sample MNHN-IK-2015-517) or lightly polysiphonic (all remaining material) stems; in addition, their hydrothecae are free from their corresponding internodes for ca ⅓ of their adaxial length, fitting the original account by Jäderholm (1903).
There is no thorough description available of the mode of branching in this species, especially at microscopic level. The original account is quite evasive (“ Die Verästelung ist sehr regelmässig und characteristisch. Die nach allen Seiten gerichteten Äste sitzen fast immer zu zweien zusammen, sind sehr deutlich spiralförmig angeordnet und nach aussen gebogen ” [The branching is very regular and characteristic. The branches are directed to all sides, almost always they are given off in pairs, are arranged in an obvious spiral, and are bent outwards]). However, Jäderholm (1903: pl. 14 fig. 5) provides a comprehensive illustration of the macroscopic appearance of a typical colony. According to his figure, a pair of plumes is given off at each geniculation of the stem, the successive pairs facing outwards with respect to the spiral built up by the stem.
Subsequent accounts seem to provide rather divergent data. Indeed, the material studied by Stechow (1909: 103, fig. 8) was devoid of the characteristic spiral growth pattern, the main stem giving rise, here and there, to secondary branches that further divide trichotomously. The colony studied by Billard (1918: 25, as Halicornaria sibogae ) formed a helicoid sympodium (“ Colonie […] ramifiée en sympode hélicoïde ”), thus partly agreeing with the type, although nothing is said about its branching pattern. The account given by Vervoort (1966: 165) (“The structure of the colony is sympodially [sic]: the main axis is formed by the basal parts of successive plumes, arranged in spiral fashion, each succeeding plume rising about 8 mm from the base of the preceding plume”) does not conform with the structure of the type either since, at each geniculation, only one plume, instead of two, is given off. Rees & Vervoort (1987) observed a “Sympodially built stem with helically arranged, unbranched plumes …” in their material, resembling those examined earlier by Vervoort (1966).
On the other hand, the colony illustrated by Rho & Park (1980: pl. 9 fig. 1) is undoubtedly similar to that of the type, despite their description ( Rho & Park 1980: 27) providing only limited details: “Colonies large, not fascicled, spirally branched. The hydrocaulus [is] divided into regular internodes, zigzagshaped”.
Finally, Vervoort & Watson (2003: 289) claim that they “have little to add to the descriptions of this characteristic species given by previous authors”.
As noted above, the branching pattern met with in the present material from New Caledonia is superficially trifid, as follows: the stem is geniculate and describes a spiral; at more or less regular intervals, the main tube bifurcates at each geniculation: in the exact continuation of the long axis of the original portion before the geniculation, it is prolonged as a plume (bearing hydrocladia), while the divergent part continues, slightly inclined (with respect to the preceding longitudinal axis), as the main stem proper; from the base of the divergent portion of the stem, two (originally twin) tubes are given off: one runs downwards along the preceding portion of the stem below the geniculation (bringing polysiphony to the colony), while the other is given away upwards, forming a second plume (bearing hydrocladia) ( Fig. 10A View Fig ).
In this context, the specific separation between G. expansum and its closest congener, Gymnangium tubulifer ( Bale, 1914) , seems to rely exclusively on the appearance of the colonies resulting from their mode of branching. When describing his species, Bale (1914) referred to the structure of the colony of G. vegae ( Jäderholm, 1903) , as did Vervoort & Watson (2003), who stated that the stem of G. tubulifer “… is geniculate and more or less spiral, composed of the proximal parts of stem elements that at each geniculation turn aside to form a branch. At the same place the proximal element splits off to form 3 ramifications of which [one] continues as the “stem”, [one] as a branch pointing away from the original element, and [one] as a downwardly directed tube, covering the more proximal parts of the “stem””. This results in a colony whose distinctive appearance is depicted by Jäderholm (1903: pl. 15 fig. 1).
In a molecular study by Ronowicz et al. (2017), G. expansum is shown to cluster with members of the genus Gymnangium Hincks, 1874 instead of with Taxella Allman, 1874 , in spite of the colony structure showing obvious affinities with the latter. As noted by these authors, Stechow (1921) proposed a new genus, Halicetta , to accommodate G. expansum , with the following diagnosis: “hydrothecae elongated, curved but not bent, with no abaxial septum”. This, however, does not differ much from characters of Taxella , even though emphasis was placed on characters of the gonosome in particular. A more comprehensive analysis is necessary to elucidate the generic affinities of Jäderholm’s (1903) hydroid.
Distribution
Scattered records from the tropical and subtropical Indian and Pacific Oceans, from Zanzibar to Japan ( Vervoort & Watson 2003).
Genus Lytocarpia Kirchenpauer, 1872
Lytocarpia brevirostris (Busk, 1852)
Fig. 11A View Fig
Lytocarpia brevirostris View in CoL – Di Camillo et al. 2011: 525, figs 2c, 3b, f, 6d, 7–8.
Material examined
PACIFIC OCEAN • several plumes branched or unbranched, 1.8–4.5 cm high, of which one bears a young corbula; off New Caledonia, stn DW4774 ; 23°01ʹ S, 168°19ʹ E; 100– 90 m; 28 Aug. 2016; KANACONO leg.; MNHN-IK-2015-514 GoogleMaps .
Remarks
For a recent description of this well-known species, refer to Di Camillo et al. (2011), who also provided an extensive synonymy.
Distribution
Widespread in tropical parts of the Indian Ocean, from Africa to Indonesia, and the northeastern South Pacific (Di Camillo et al. 2001).


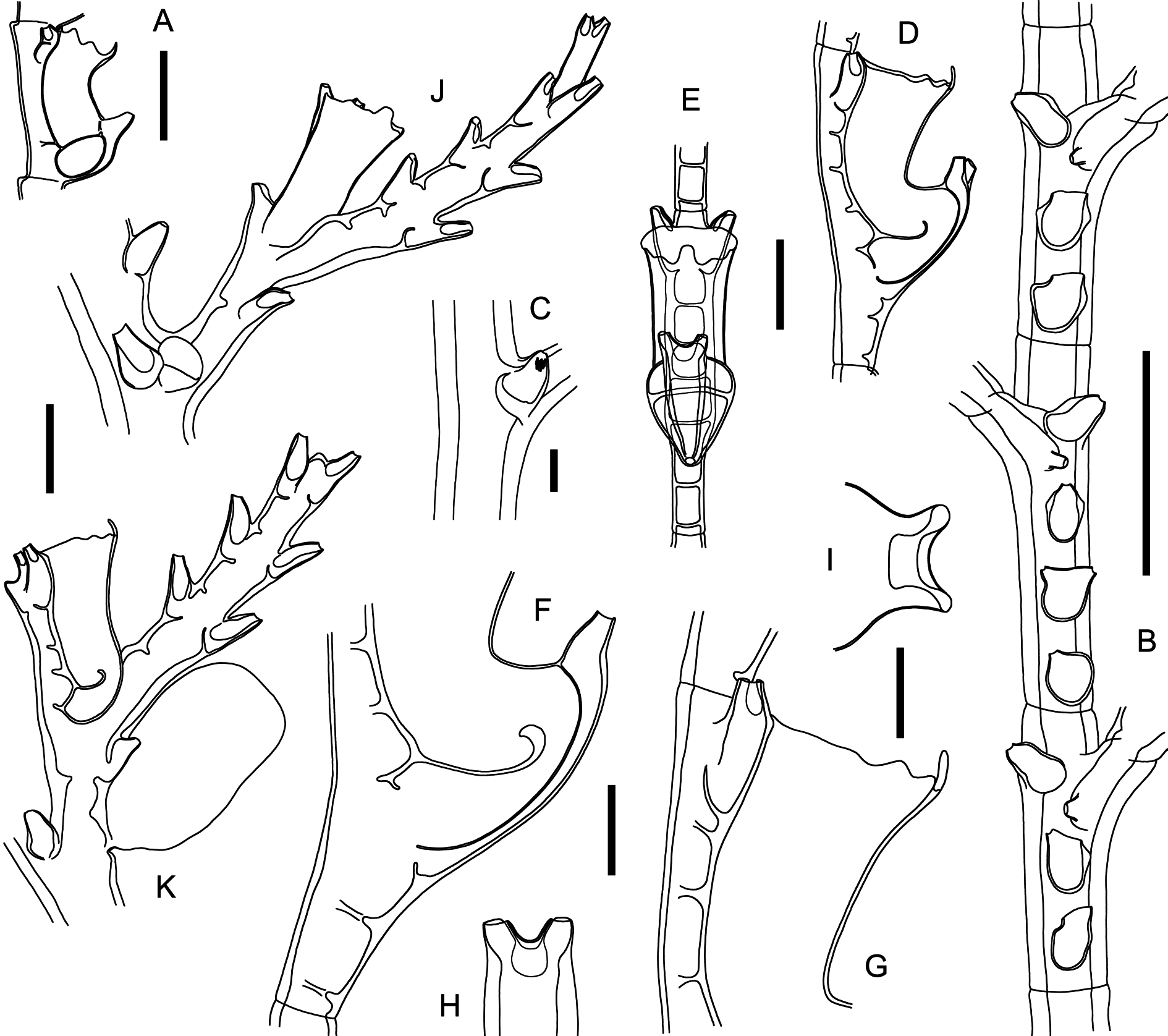
Table 2. Cladocarpus-like hydroids with S-shaped hydrothecae provided with an adaxial septum, and their distinctive features with respect to Cladocarpus pennatus sp. nov.
| Nominal species | Brief description, with emphasis on the distinctive features, and geographical distribution |
|---|---|
| Stem internodes with 1 frontal nematotheca and 2 axillar, flanking each cladial apophysis on both | |
| Cladocarpus cartieri Bedot, 1921 | sides of the stem; no mamelon; cormidia long, slender, with up to 9 intranodal septa, one of which is proximal and abaxial; hydrothecae elongated, adaxial septum short, aperture rounded, rim entirely smooth; mesial nematotheca a short distance below hydrothecal base, laterals elongate, overtopping aperture ( Ramil & Vervoort 1992b). Distribution: Azores, Mid Atlantic Ridge (Calder &Vervoort |
| 1998). | |
| Streptocaulus dollfusi (Billard, 1924) | Stem internodes with 2–5 frontal nematothecae, a fronto-axillar one, but no conical nematotheca on cladial apophysis; cormidia with 6–8 intranodal ridges; hydrothecae elongated, adaxial septum set upwards, rim with 1 median abaxial cusp variably developed, elsewhere smooth to weakly undulated; mesial nematotheca below hydrothecal base, laterals facing upwards and forwards, overtopping hydrothecal rim; phylactocarps borne laterally on 1 st hydrotheca, composed of a succession of short internodes with distal pairs of nematothecae ( Ansín Agís et al. 2001); gonothecae on phylactocarps: female elongated, aperture lateral, subterminal, male fusiform, distally truncate ( Billard 1934). |
| Distribution: eastern Atlantic from the Bay of Biscay to Morocco ( Ansín Agís et al. 2001). | |
| Stem with 2–4 frontal nematothecae (each with 4 apertures) between successive cladial apophyses, single axillar nematotheca, and no conical nematotheca on apophysis; cormidia long, S-shaped, Cladocarpus with centrally-placed hydrotheca; mesial nematotheca with 4 apertures, seated below the millardae hydrothecal base; lateral nematothecae antler-shaped, with 5–7 apertures; hydrothecal rim without Vervoort, 1966 cusps; internodes of phylactocarp with 2 lateral nematothecae, each with 2–3 apertures; gonotheca globular, distally truncate. Distribution: Mozambique, NE coast of South Africa ( Millard 1975). | |
| Cladocarpus paraventricosus Ramil & Vervoort, 1992 | Stem internodes with 1 frontal nematotheca, single axillar nematotheca, and no conical nematotheca on apophysis; cormidia long, slightly sigmoid, with centrally-placed hydrotheca; mesial nematotheca below the hydrothecal base, laterals long, tubular, surpassing the rim; margin with median, conspicuous cusp and 5 pairs of laterals, slightly marked; intrathecal septum short, projecting upwards; phylactocarps borne on first cormidium, branched pseudodichotomously, a gonotheca after each ramification; gonotheca ovoid, aperture lateral, subapical. Distribution: Strait of Gibraltar ( Ramil & Vervoort 1992b). |
| Stem with 1–5 frontal nematothecae between successive cladial apophyses, with single axillar | |
| nematotheca, and no conical nematotheca on apophysis; cormidia long, slightly sinusoid, Cladocarpus hydrotheca placed in middle portion, elongated, mesial nematotheca below the hydrothecal base; sinuosus rim of hydrotheca, besides the median abaxial cusps, with faintly sinuous lateral edges; internodes Vervoort, 1966 of phylactocarp with two very long, curved nematothecae; gonothecae ovoid with truncate distal end ( Vervoort 1966; Millard 1975). Distribution: Alboran Sea, Guinea Bissau, South Africa (Ansín Agís et al. 2001). | |
| Main tube of the stem with intranodal septa, 1–3 frontal nematothecae between successive cladial Cladocarpus apophyses, single axillar nematotheca, and no conical nematotheca on apophysis; hydrotheca unicornus with basal abaxial bulge capped by short solid horn of perisarc; mesial nematotheca free from Millard, 1975 hydrotheca, lateral nematothecae elongated, with 3 apertures, margin with median, abaxial, inturned cusp, elsewhere smooth. Distribution: endemic to South Africa ( Millard 1975). | |
| Stem internodes with 3–8 frontal nematothecae, 2 axillar nematothecae, and a mamelon on each hydrocladial apophysis; cormidia fairly long, with up to 9 intranodal septa; hydrotheca ventricose, Cladocarpus adaxial ridge directed upwards; mesial nematotheca below the hydrothecal base or slightly surpassing ventricosus it; lateral nematothecae overtopping hydrothecal rim; rim with median abaxial cusp and slightly Allman, 1877 crenate edges; gonothecae borne on stem, oval, aperture lateral, subterminal; phylactocarp borne on first cormidium, antler-shaped, with 3–4 branchlets ( Bogle 1975; Ramil & Vervoort 1992b). Distribution: Florida and Straits of Florida ( Bogle 1975). | |
| Stem with at least one frontal nematotheca, a pair of axillar nematothecae, and a mamelon; internodes as long as to accommodate a hydrotheca and its 3 nematothecae; hydrotheca fairly long, Cladocarpoides adaxial septum projecting upwards, mesial nematotheca adnate to lower abaxial thecal wall, laterals yucatanicus overtopping margin; rim sinuous, except for the median, abaxial cusp; gonothecae obovate, borne on Bogle, 1984 base of stem apophyses; phylactocarp borne on first cormidium, corbula-like, costae antler-shaped, with associated hydrotheca and terminated in nematophorous spike. Distribution: Yucatan Channel ( Bogle 1984). | |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Hydroidolina |
|
Order |
|
|
Family |
|
|
Genus |
Cladocarpus pennatus
| Galea, Horia R. 2020 |
Lytocarpia brevirostris
| Di Camillo C. G. & Puce S. & Bavestrello G. 2011: 525 |
Gymnangium expansum
| Rees W. J. & Vervoort W. 1987: 163 |
| Vervoort W. 1966: 165 |