
Oligosoma toka, Chapple, David G., Bell, Trent P., Chapple, Stephanie N. J., Miller, Kimberly A., Daugherty, Charles H. & Patterson, Geoff B., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.205462 |
|
DOI |
https://doi.org/10.5281/zenodo.6182746 |
|
persistent identifier |
https://treatment.plazi.org/id/03AC4C11-8D37-A42D-FF4C-FB7DFF064A69 |
|
treatment provided by |
Plazi |
|
scientific name |
Oligosoma toka |
| status |
sp. nov. |
Oligosoma toka sp. nov.
Figure 8 View FIGURE 8. a
Oligosoma inconspicuum Jewell 2008: 88
Leiolopisma inconspicuum Patterson & Daugherty 1990: 66
Holotype. Schoolhouse Flat, Nevis Valley, (45º 11’S, 168º 59’E), RE007278, adult male (coll. T. Bell, 2009).
Paratypes (12 specimens). Nevis Range F42 5551813.9 2183702.9 (45º 10’S, 168º 52’E), 7 specimens (RE006165 [CD938], female; RE006166 [CD939], male; RE006163 [CD936], female; RE006164 [CD937], female; RE006167 [CD940], male; RE006168 [CD941], female; RE006162 [CD935], female) (coll. A.H. Whitaker, March 1986); Schoolhouse Flat, Nevis Valley, (45º 11’S, 168º 59’E), 5 specimens (RE007281, female; RE007286, female; RE007289, female; RE007290, male; RE007297, male) (coll. J. Reardon January 2010).
Live animals examined. Schoolhouse Flat, Nevis Valley (45º 11’S, 168º 59’E), 13 specimens (5 adult males, 5 adult females, 3 juveniles) (data collected by T. Bell, 2009).
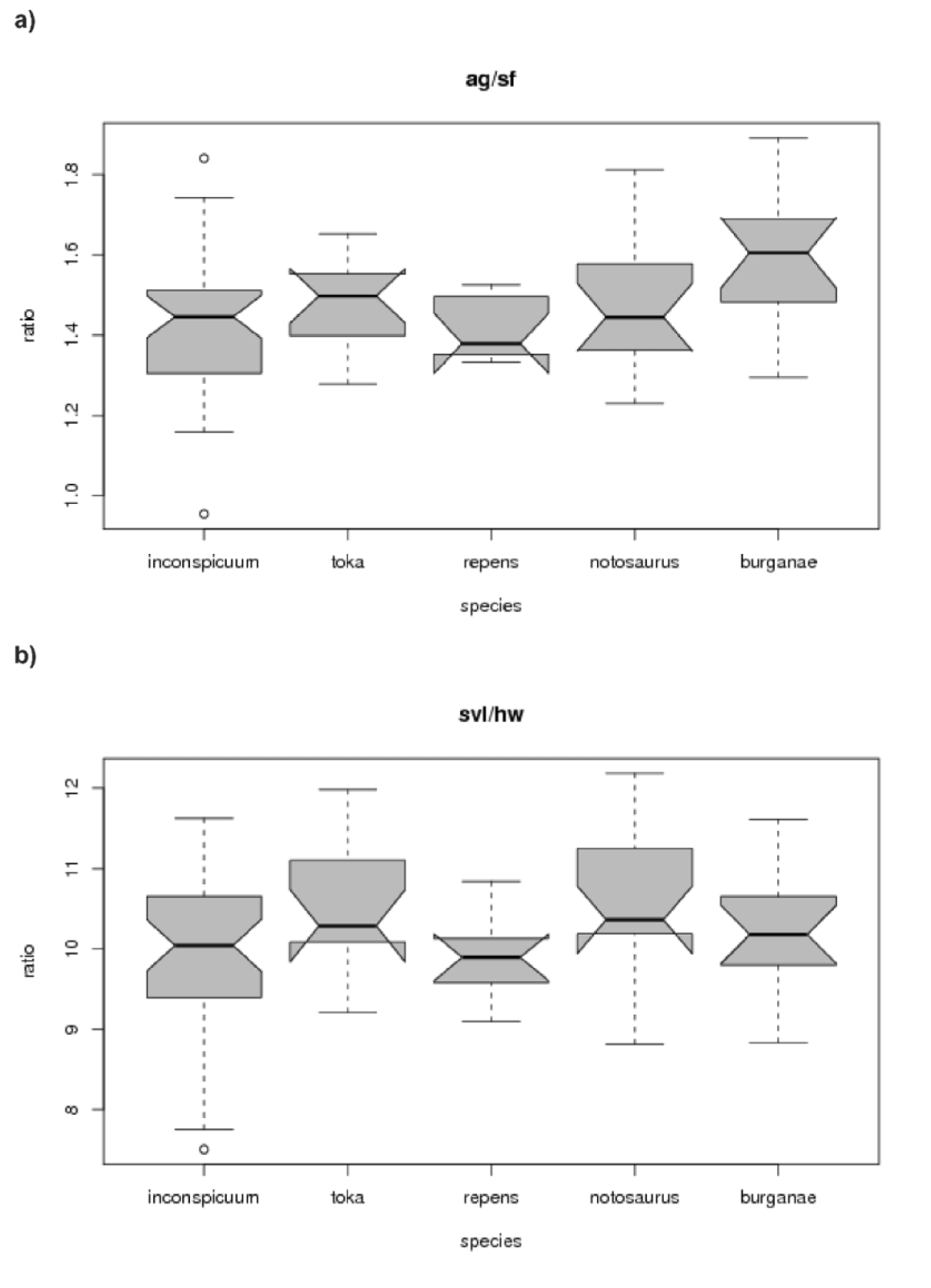
Diagnosis. Oligosoma toka can be distinguished from other related Oligosoma species through a combination of characters ( Figure 4 View FIGURE 4 a – b ). Compared to O. maccanni , O. toka has a glossy appearance, with brown predominating whereas O. maccanni has a greyer ground colour. Oligosoma maccanni has a pale grey ventral colour rather than the yellow ventral colour seen in O. toka . The ear opening in O. maccanni often has large projecting scales on the interior margin, whereas these are often minimal or lacking altogether in O. toka . Oligosoma maccanni has four supraocular scales compared with three in O. toka , an unusually low number for New Zealand skinks. Sympatric O. polychroma have very similar colour patterns, but can be distinguished by a pale dorsal stripe on the outside of the forelimbs, and a greyish-brown ventral colouration. The ear opening in O. polychroma often has prominent projecting scales on the interior margin. There are statistical differences between O. toka and O. repens sp. nov. (SVL/HL, SVL/HLL, ventral scales, SE/EF), O. burganae (SVL/HLL, ventral scales), O. inconspicuum (SVL/FL, SVL/HLL, ventral scales), and O. notosaurus (ventral scales) ( Figure 4 View FIGURE 4 a – b ). All O. toka have three supraoculars whereas all O. inconspicuum and O. notosaurus have four. The number of ventral scales in O. tekakahu (68) is fewer than O. toka (70–88), and the number of subdigital lamellae (16) is fewer than O. toka (17–23). The dorsal surface of the head is usually more strongly marked than in O. repens sp. nov., and the mid-dorsal and dorsolateral stripes in O. toka are more prominent than in O. repens sp. nov..
Description of Holotype. Body elongate, oval in cross-section; limbs moderately well-developed, pentadactyl. Lower eyelid with a transparent palpebral disc, bordered on sides and below by small, oblong granules. Nostril centred just below middle of nasal, pointing up and back, not touching bottom edge of nasal. Supranasals absent. Rostral broader than deep. Frontonasal broader than long, not separated from frontal by prefrontals meeting in midline. Frontal longer than broad, shorter than frontoparietal and interparietal together, in contact with 2 anteriormost supraoculars. Supraoculars 3, the second is the largest. Frontoparietals distinct, larger than interparietal. A pair of parietals meeting behind interparietal and bordered posteriorly by a pair each of nuchals and temporals, also in contact with interparietal, frontoparietal, third supraocular and 2 postoculars. Loreals 2, similar size; anterior loreal in contact with first and second supralabial, posterior loreal, prefrontal, frontonasal and nasal; posterior loreal in contact with second supralabial, first subocular, upper and lower preocular, prefrontal and anterior loreal. Supralabials 8[left]/7[right], the sixth and seventh are the equal largest. Infralabials 6, several of them equal in size; sixth[left]/ fifth[right] supralabial below centre of eye. Mental broader but shallower than rostral. Suboculars 3 and 4 separated by sixth[left]/fifth[right] supralabial. Chinshields 3 pairs. One primary temporal, similar size to lower secondary temporal. Dorsal scales similar in size to ventral scales, weakly striate. Ventral scales smooth. Subdigital lamellae smooth. Ear opening round, small with insignificant projecting granules. Forelimbs shorter than hindlimbs. Adpressed limbs not meeting in adult. Digits moderately long, sub-cylindrical. Third front digit shorter than the fourth.
Measurements (in mm; holotype with the variation shown in the type series in parentheses). SVL 71.1 (mean 59.8, range 45.1–66.2), HL 8.8 (mean 8.2, range 6.9–9.0), HW 6.4 (mean 5.7, range 4.8–6.5), AG 40.5 (mean 32.0, range 24.1–40.5), SF 24.5 (mean 21.7, range 17.0–24.5), SE 12.2 (mean 10.0, range 8.7–12.2), EF 12.6 (mean 11.7, range 8.3–13.8), and TL 58.5 (mean 58.5, range 53.5–66.0, N=3).
Variation (holotype with the variation shown in the type series in parentheses). Upper ciliaries 7 (mean 6, range 5–7); lower ciliaries 8 (mean 8, range 7–9); nuchals 4 pairs (mean 3 pairs, range 2–4 pairs); midbody scale rows 32 (mean 32, range 30–34); ventral scale rows 80 (mean 82, range 70–88); subdigital lamellae 21 (mean 21, range 17–23); supraciliaries 5 (mean 5, range 5–7); suboculars 7 (mean 6, range 4–7). Frontonasal seldom separated from frontal by prefrontals meeting in midline. Anterior loreal in contact with first or second supralabial. Secondary loreal usually in contact with secondary supralabial only. Supralabials 6, 7 (usual) or 8, the fifth, sixth or seventh are the largest. Infralabials 5, 6 (usual) or 7. Third front digit as long as (usual) or shorter than the fourth. Maximum SVL 71.1 mm. Three specimens had an intact tail (TL/SVL = 1.16). Ratios for morphological measurements (± SD): AG/SF 1.47 ± 0.12; SE/EF 0.86 ± 0.11; HL/HW 1.45 ± 0.06.
Colouration. Dorsal surface light to dark yellowish brown, often with irregular flecks. A median dorsal dark yellowish brown longitudinal stripe, 2 half-scale rows wide, well developed, commencing behind the head and passing back to the base of the tail, becoming indistinct thereafter. A light to grayish yellowish brown dorsal band 2 half-scale rows wide sometimes with light and dark flecks. This band is often bounded on both sides by a pale stripe less than one scale wide. Another dark yellowish brown band 1 to 2 half-scale rows wide, shading onto a prominent pale dorsolateral band 2 half-scale rows wide. This pale band extends from above posterior margin of eye to base of tail, or further along tail. A broad strong yellowish brown lateral band 1 to 2 scale rows wide, originating near tip of snout, passing through eye and ending at base or further along tail, bordered laterally by two dark yellowish brown bands, and often with pale scales extending into it from above and below; sometimes flecked with white. Below this an indistinct pale stripe passes from beneath the posterior border of the eye above the ear and limb insertions to the tail. This stripe is irregularly defined below by brown scales which merge gradually with the yellow ventral colouration. Yellow colouration extends along first third of tail. Ventral surface usually lightly speckled with black spots on chin and throat, which are white. Outer surface of forelimbs is dark brown with black and white specks. Juvenile colouration similar to adult, but generally lighter and lacking distinct mid-dorsal stripe. There do not appear to be sexually dimorphic colour patterns.
Etymology. From ‘toka’, the Maori word for rock or boulder. Refers to the rocky habitat on which this species occurs in the Nevis Valley. The common name is the Nevis skink.
Habitat and life history. Oligosoma toka appears to be confined to Nevis Valley (WAIKAIA 74.01 Nokomai Ecological District; McEwen 1987) of central Otago ( Figure 5 View FIGURE 5 e). The area where O. toka has been recorded so far is classified as Environments N3, N4 and Q3 ( Leathwick et al. 2003). The Nokomai ED consists of broad plateaus and hills of a lower altitude (> 600 m to 1500 m asl) than the Eyres ED. The geological composition is complex, but usually consists of schist and greywacke rock formations. The climate is cool, with annual rainfall typically around 800–1200 m. It is unclear how far south the Nevis skink population extends to, but it is likely to be the entire valley and adjacent areas. The predominant ecology consists of exotic pasture for sheep and cattle grazing, with lowland tussockland in the foothills, and red or subalpine tall tussockland at higher elevations. Oligosoma toka are extremely abundant around rock piles (old gold tailings) along the eastern side of the Nevis River, but not as abundant elsewhere where cover is scarce on the open Nevis Valley flats. However, they do occur widely in the area, on the river flats, around the foothills and up to the Nevis Crossing. The artificial rock piles are likely to be important refugia for the population in the Nevis Valley, and O. toka is the most abundant species in the valley, especially adjacent to the Nevis River (T. Jewell, pers. obs.). However, high abundance at certain localities (e.g. 5– 7 skinks per 10 m 2 in artificial and natural rock formations; T. Jewell, unpublished data) does not necessarily imply overall species security, given the apparent restricted range of the species. Important vegetation for O. toka include tussocks and rank grasses (native, exotic), Coprosma , Discaria Muehlenbeckia , Melicytus and Rubus spp. In the wider Nevis Valley, O. toka is sympatric with O. inconspicuum , O. polychroma , O. maccanni , Hoplodactylus sp. ‘Cromwell’, and Hoplodactylus sp. ‘Otago large’ (T. Jewell, pers. obs.; this study). It is possible that O. toka might also be present in the adjacent Hector and Garvie Mountains (T. Jewell, pers. comm.).
Conservation status. Little is known about the range, abundance and population viability of O. toka . It is currently considered Data Deficient in the New Zealand Department of Conservation’s national threat classification lists ( Hitchmough et al. 2010). Resolution of this species’ conservation status may be urgent ( Townsend et al. 2008).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Oligosoma toka
| Chapple, David G., Bell, Trent P., Chapple, Stephanie N. J., Miller, Kimberly A., Daugherty, Charles H. & Patterson, Geoff B. 2011 |
Oligosoma inconspicuum
| Jewell 2008: 88 |
Leiolopisma inconspicuum
| Patterson & Daugherty 1990: 66 |