
Discocelis hollemani, Maghsoudlou, Abdolvahab & Rahimian, Hassan, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3683.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:845A5A13-A77B-4181-8CAB-D2D4170954DF |
|
DOI |
https://doi.org/10.5281/zenodo.6157880 |
|
persistent identifier |
https://treatment.plazi.org/id/8E5BC00E-AFEB-431D-89DA-1998FD473FCD |
|
taxon LSID |
lsid:zoobank.org:act:8E5BC00E-AFEB-431D-89DA-1998FD473FCD |
|
treatment provided by |
Plazi |
|
scientific name |
Discocelis hollemani |
| status |
sp. nov. |
Discocelis hollemani View in CoL sp. nov.
( Figures. 7–11 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 )
Material examined and locality. Holotype: one mature specimen with the anterior half mounted on glass slide and the posterior half, containing the copulatory apparatus, as series of sagittal sections (Six slides, ZUTC Platy.1249).
Paratypes: three mature sectioned specimens with the anterior half of each mounted on a microscope slide and the posterior half of each, containing the copulatory apparatus, as series of sagittal sections (six slides per specimen) ( ZUTC Platy.1250, 1251, 1252); one mature specimen cleared and stained with alum carmin ( ZUTC Platy.1253); two mature specimens preserved in ETOH 70% ( ZUTC Platy.1254).
Etymology. The specific epithet is dedicated to Prof. John Holleman, who has been working extensively of Acotylean and Cotylean polyclads and was very helpful during the course of the present study.
Type locality. All specimens were found together, in March 2011, under a rock, during the low tide, from the rocky shores of Chapahn station, the Persian Gulf, 28° 20´47ʺ N, 51° 10´59.8ʺ E ( Fig. 1 View FIGURE 1 , station 2).
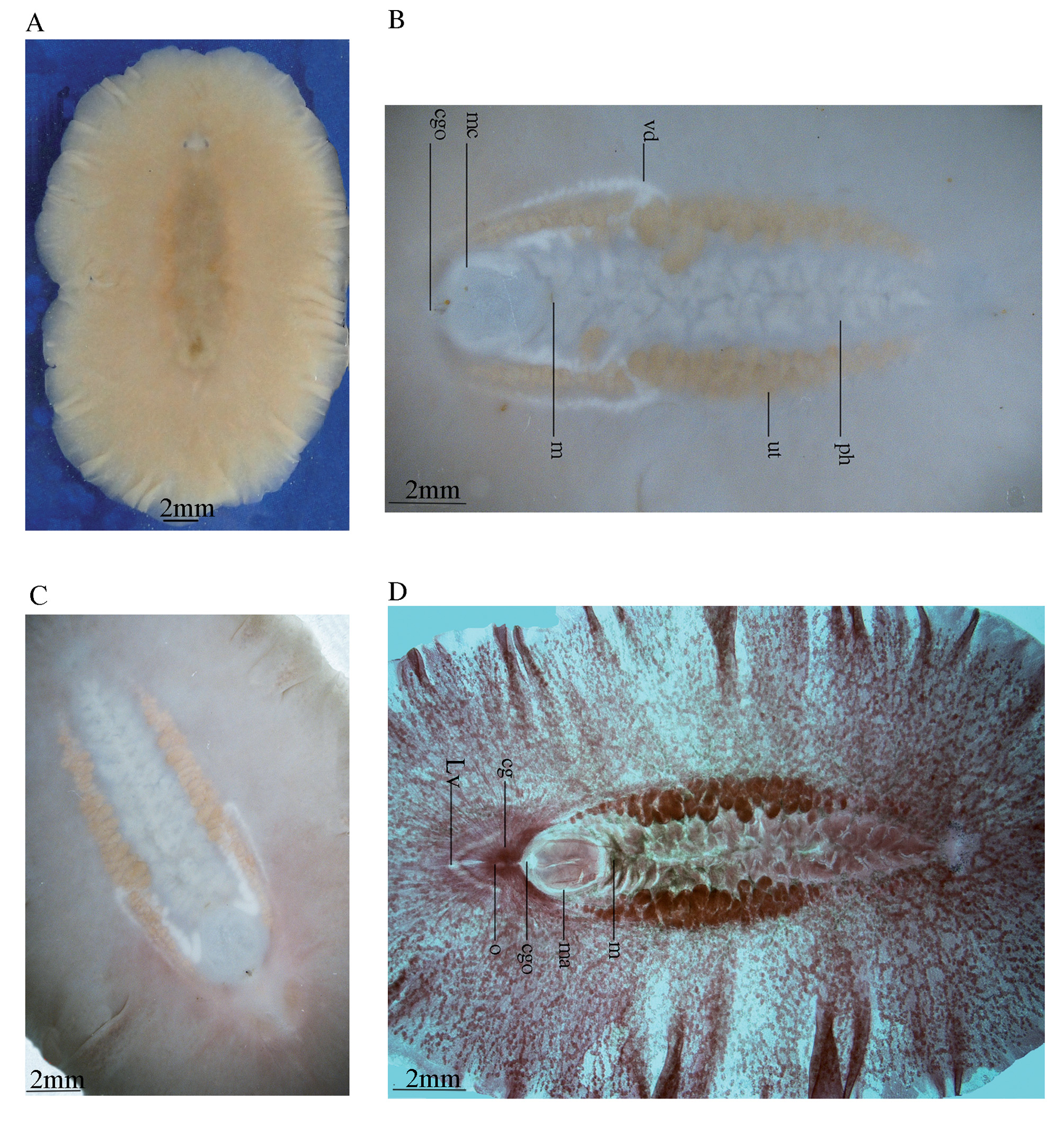
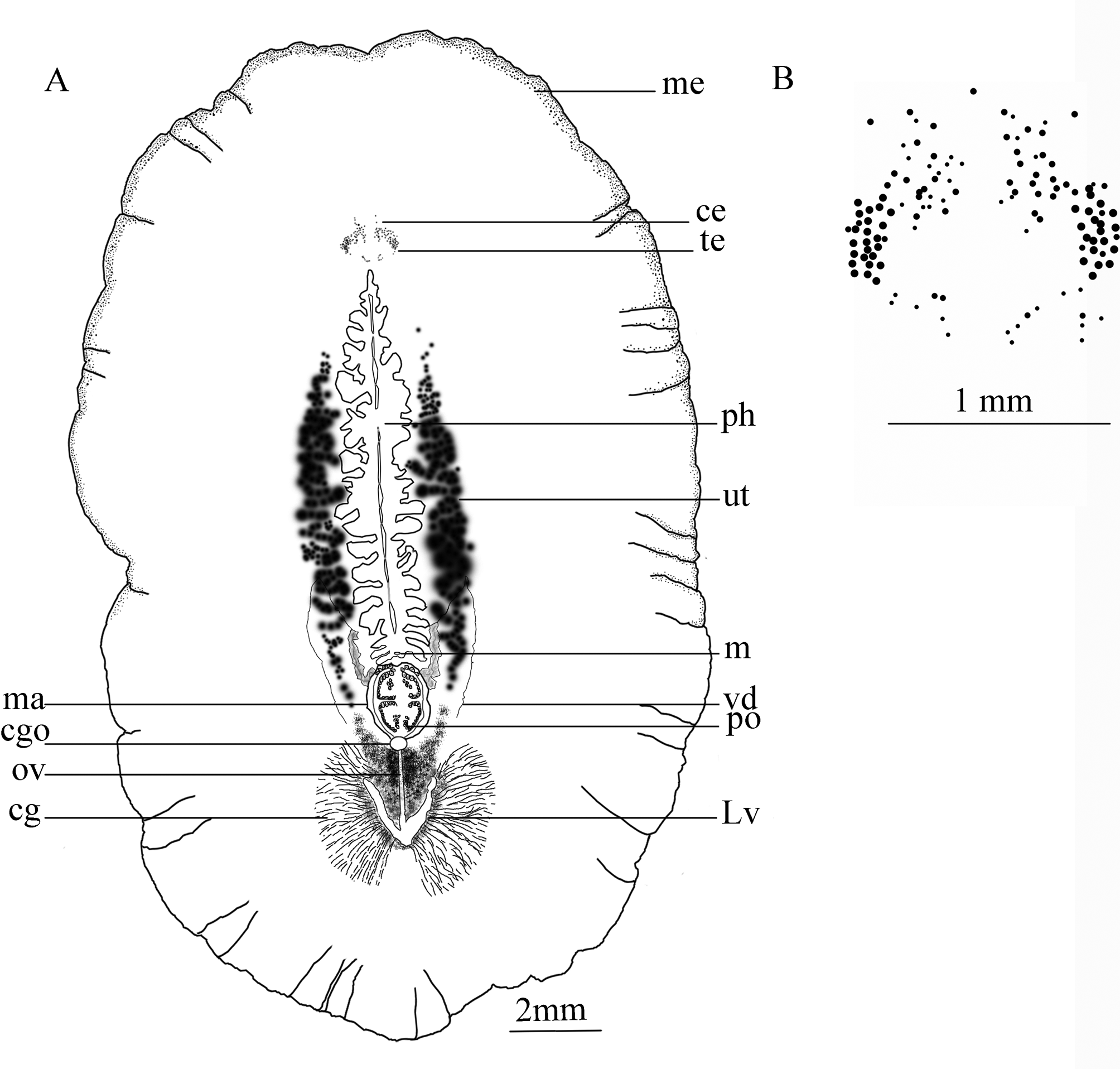
Description. External morphology. Fixed specimens measured from 3.5 × 2 cm in the holotype, to 4 × 2.2 (SD±0.54×0.25) cm in the paratype series (n=7); body broadly oval, thick and with a fleshy consistency. Dorsal surface of living worms light yellowish-brown to wheaten, without any spots, shading to brown mid-dorsally, with translucent margins ( Fig. 7 View FIGURE 7 A); ventral surface beige in the holotype ( Fig. 7 View FIGURE 7 B), mistyrose in some paratype individuals, fading towards the margin and becoming light pink around the common gonopore ( Fig. 7 View FIGURE 7 C). Orange uterus prominent, appearing as a cluster round the pharynx ( Fig. 7 View FIGURE 7 B, in holotype and Figs, 7C–D in paratypes). No tentacle present. Tentacular eyes visible against a white background ( Fig. 7 View FIGURE 7 A), as two converge half-crescentic clusters, with 25–27 eyespots in each cluster ( Fig. 8 View FIGURE 8 B), each 0.4 mm long, 1.2 mm apart; cerebral eyes embedded in the epidermis, and more difficult to observe in the living animal, but readily observable in a cleared paratype specimen (ZUTC platy 1253; Fig. 7 View FIGURE 7 D), two clusters each subdivided into anterior and posterior groups in relation to the cerebral organ: Anterior group extend to the length half of the distance between the two tentacular eyes (0.6mm), with 20–25 eyespots in each cluster; posterior group with 8–10 eyespots in each cluster; a band of small unequal eyes, in four to five rows, about 0.4 mm thick, extend on the margin to near the posterior end of pharynx ( Fig. 8 View FIGURE 8 A).
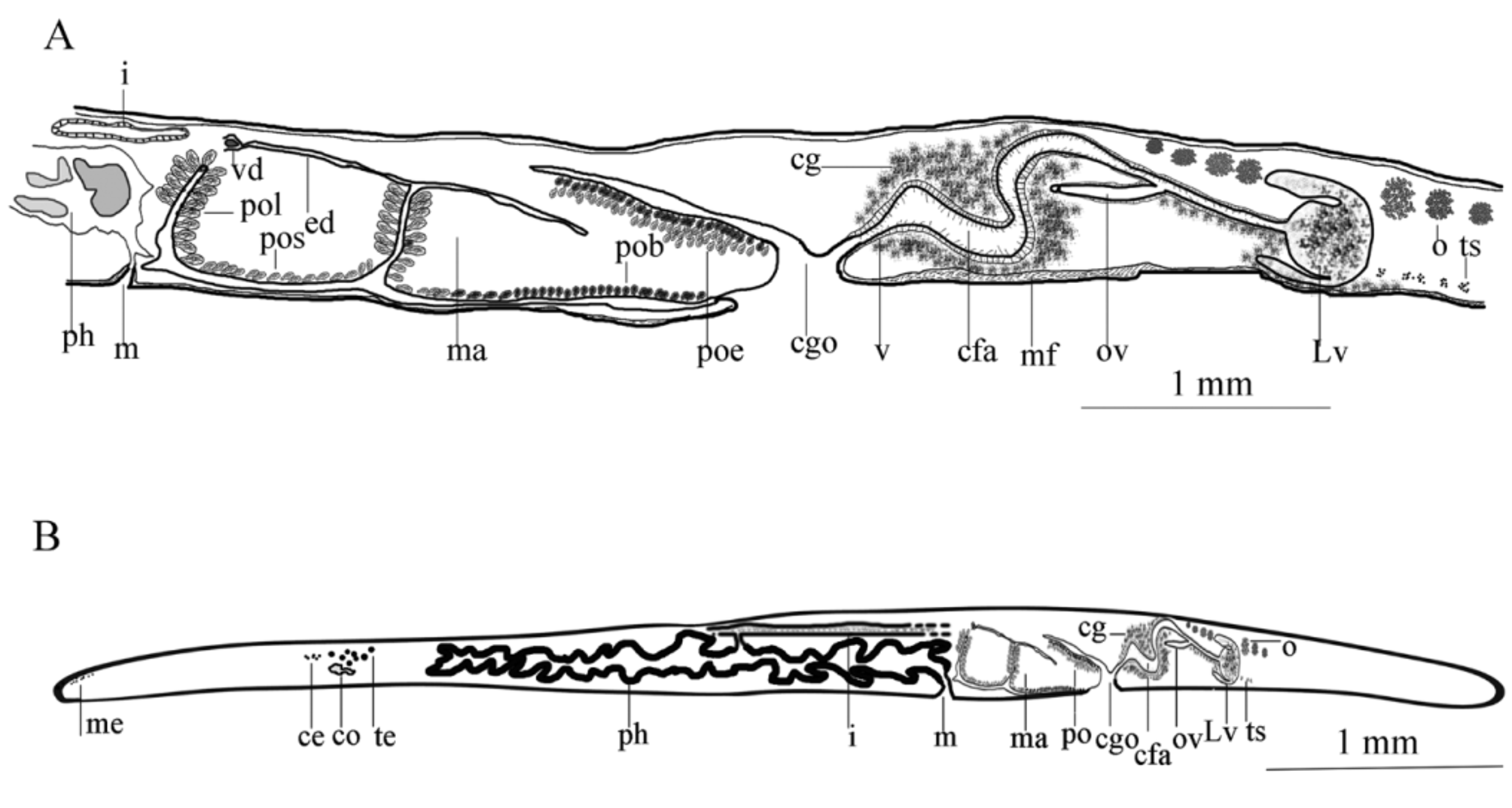
Digestive system: Ruffled pharynx with 12 to 13 pairs of folds, situated in the central part of the body, become somewhat larger posteriorly, begins immediately posterior to the cerebral eye clusters, 5 mm from the anterior end and 12.8 mm from posterior end of the body; pharynx 9.8 mm long, about one-third of the body length, width of pharynx 2.5 mm at the midpoint, the last posterior fold of the pharynx extends posterio-ventrally to the male complex; mouth located at the posterior end of the pharyngeal cavity, 13.3 mm from posterior end, immediately behind the mouth is the massive male complex ( Figs. 7 View FIGURE 7 B, 8A).
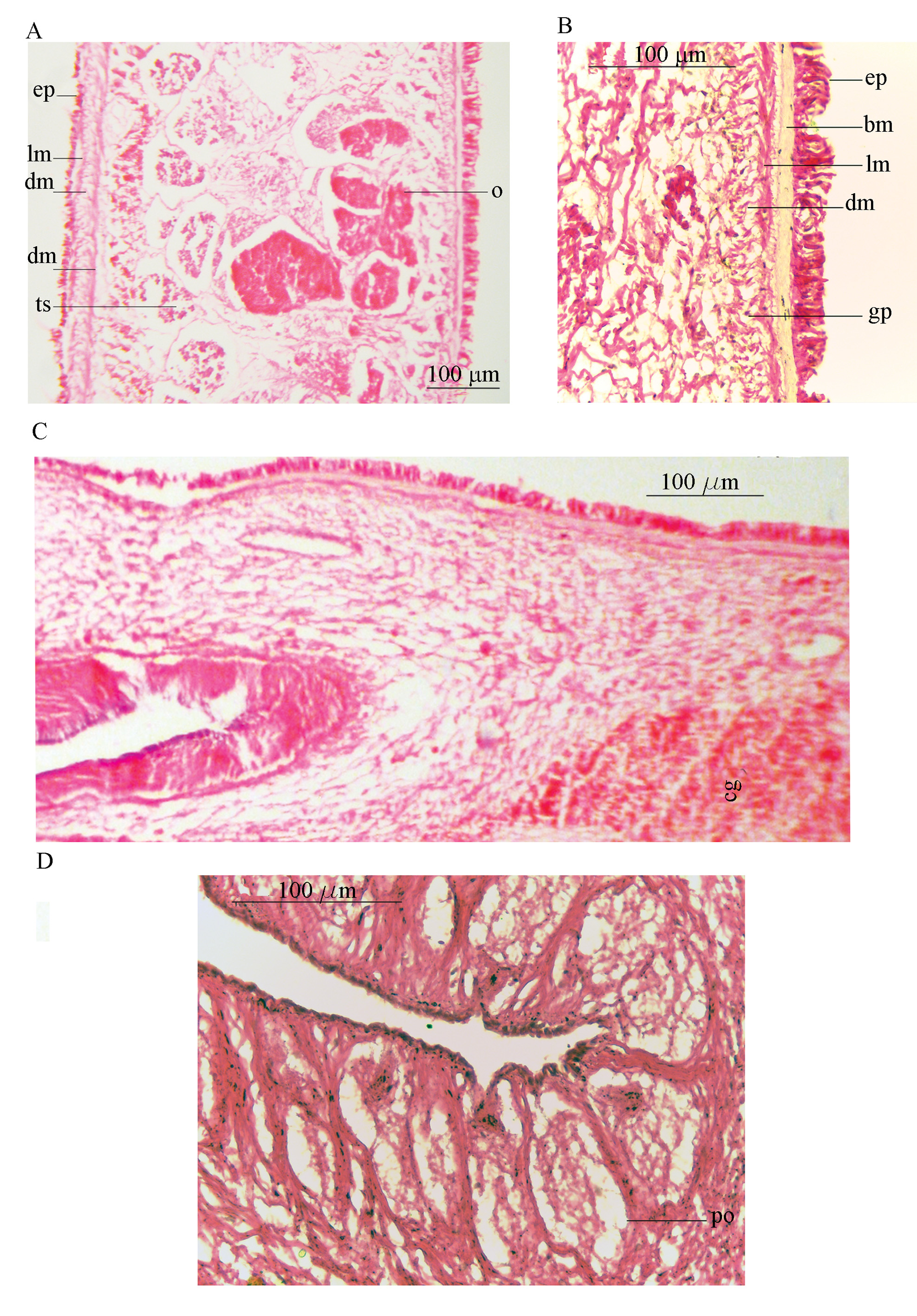
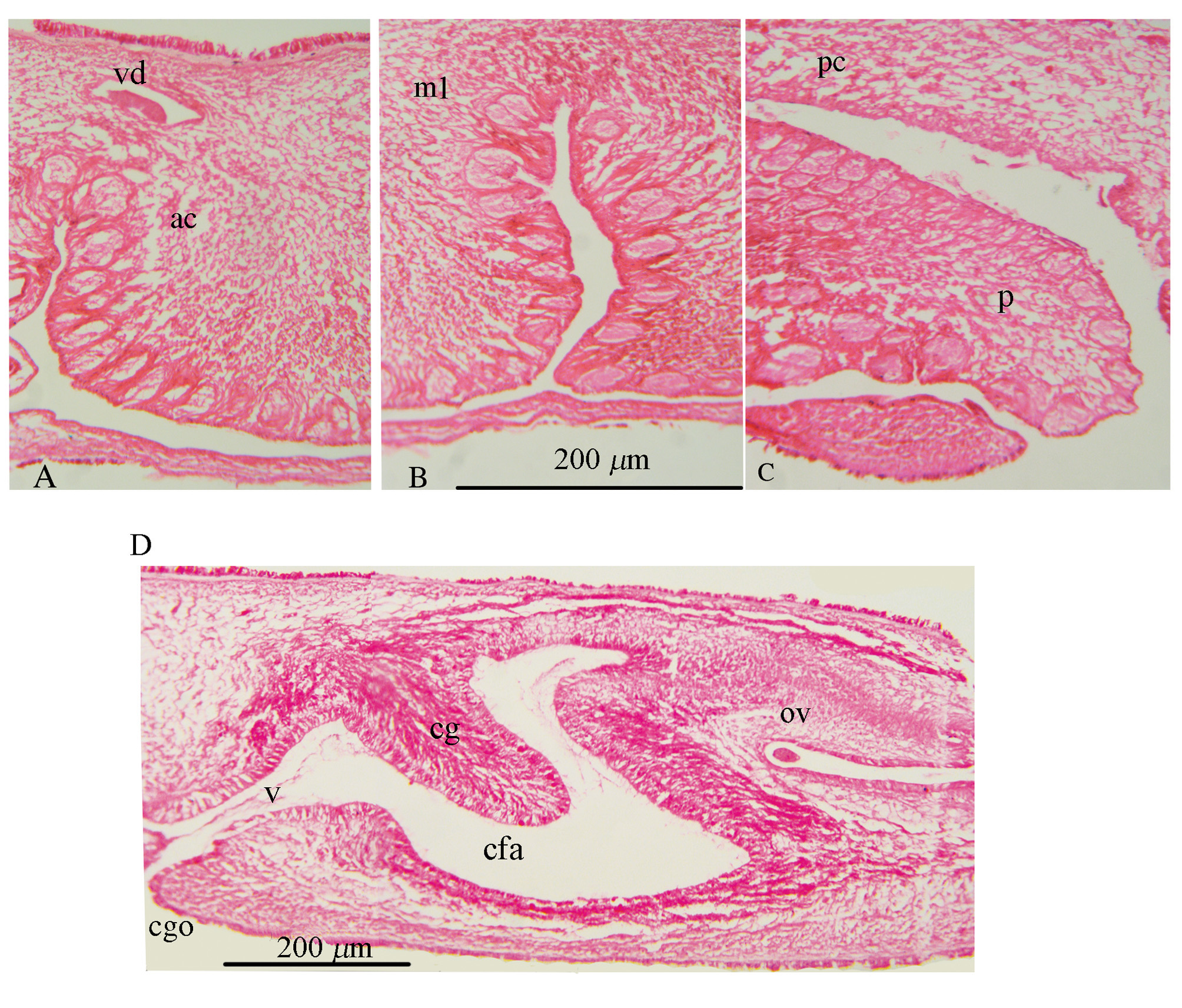
Epidermis and body wall: Ventral body wall thickness about 43 µm at the anterior region ( Fig. 9 View FIGURE 9 A), 28 µm around the male complex, and becomes thicker around the female system (86 µm) ( Fig. 9 View FIGURE 9 C); epidermis without rhabdites, composed of three layers: a longitudinal muscle layer (about 10 µm), a well-developed layer of mixed circular and diagonal muscle fibers (about 23 µm), and the innermost longitudinal muscular fibers merging with diagonal ones (11µm). Beneath the muscular layer, in the parenchyma, distinct basophil glandular cells are distributed among numerous transverse muscle fibers ( Fig. 9 View FIGURE 9 A).
Dorsal body wall 37 µm thick, with ciliated epidermis, intra-epithelial nuclei and numerous rhabdites; body wall musculature consisting of a longitudinal layer (8 µm), and a combination of circular and diagonal muscle fibers (24 µm) ( Fig. 9 View FIGURE 9 B).
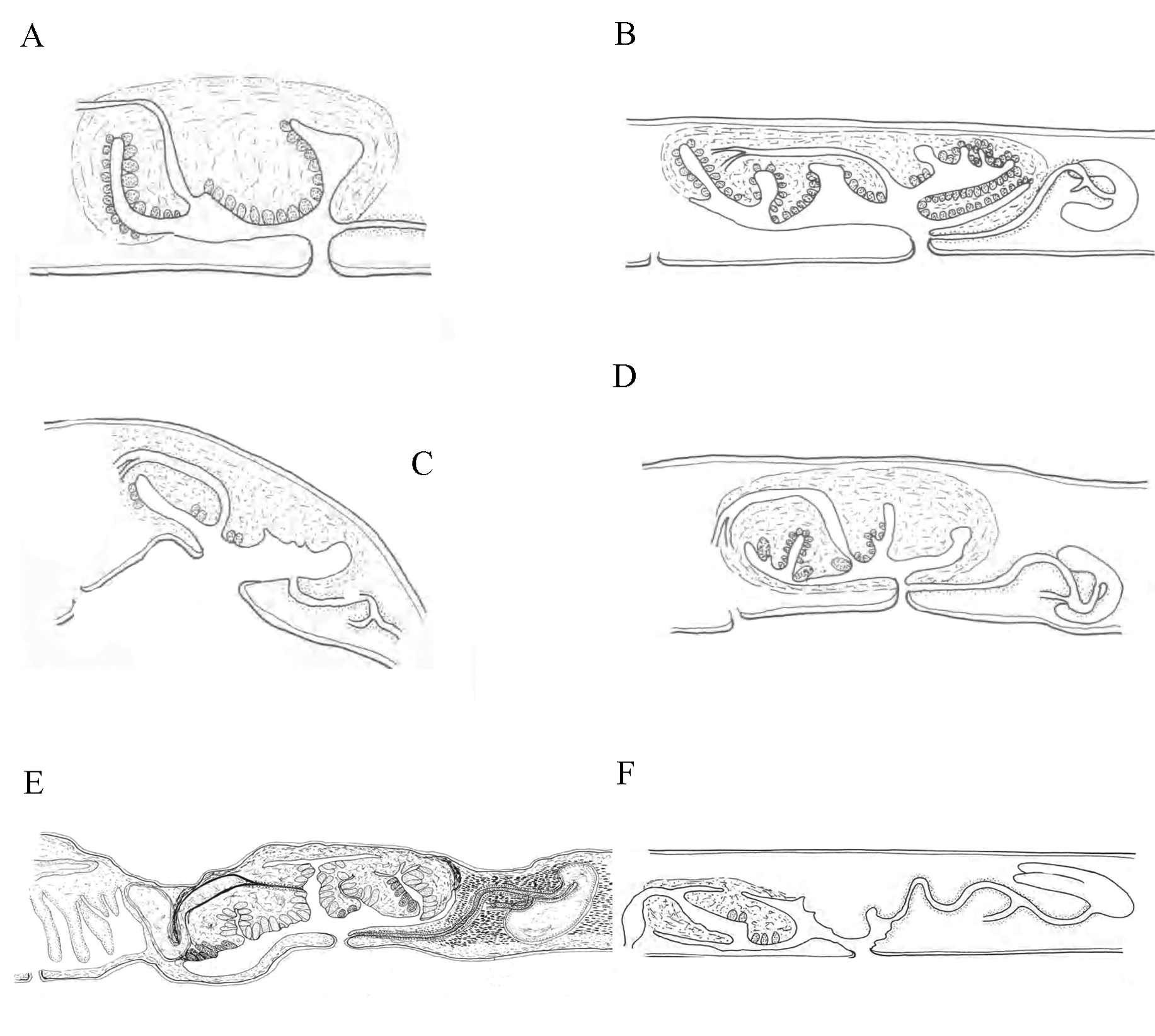
Male copulatory system: The male copulatory apparatus situated immediately behind the pharynx ( Figs. 7 View FIGURE 7 B, 8A), the spermiducal vesicle was not seen. The male atrium is bi-lobed, round in ventral view, containing a muscular lobed penis and prostatoid organs ( Fig. 8 View FIGURE 8 A). In sagittal views, the male atrium is long, somewhat bowshaped, without muscular folds, with two lobes: One anterior lobe, somehow wide, descending dorsally below the pharynx, has large prostatoid organs on dorso-ventral sides of the anterior tip ( Figs. 10 View FIGURE 10 A and 11A), the first lobe of the male atrium does not reach the mouth dorsally in sagittal views ( Figs. 10 View FIGURE 10 A, B); a longer posterior lobe, with small prostatoid organs, in two or three layers, encircle the lobe ( Figs. 10 View FIGURE 10 A, and 11C). Behind the penis the male atrium opens by a slanting route to the common gonopore. The distance between the anterior lobe and the common gonopore is 2.2 mm. Prostatoids are arranged distally in a crescent cluster on penis lobes, occur in two distinct sizes and contents: the large prostatoids of dorso-lateral sides of the anterior lobe having a faintly eosinophilic content, average size about 75× 48 µm ( Fig. 10 View FIGURE 10 A); the smaller prostatoids, especially those distributed distally on posterior lobe, are darker having a basophilic content ( Fig. 10 View FIGURE 10 B). Tear-shaped prostatoid organs filled with fine darkly staining matrix and nuclei scattered through the matrix, surrounded with muscle fibers ( Fig. 9 View FIGURE 9 D), and with narrow distal ends pass through the epithelium and empty into the male atrium; numerous extra vesicular glands are scattered around the prostatoid organs.
Vasa deferentia visible in ventral view, parallel posterior to the pharynx, extending anteriorly, up to the level of the posterior third of the pharynx, then recurrent posteriorly and dorsally to connect to the male atrium ( Figs. 7 View FIGURE 7 B, 8A). In sagittal view, the vas deferens runs posteriorly narrowing constantly, eventually entering the ejaculatory duct, in the middle of lobed penis ( Fig. 10 View FIGURE 10 A); without spermiducal vesicle. The posterior free end of penis devoid of prostatoids centrally. ( Figs. 10 View FIGURE 10 A and 11C); the male atrium, near its posterior end, opens into 0.1 mm wide common gonopore. The common genital atrium located at the anterior end of the posterior third of the body.
Female reproductive apparatus. The female reproductive apparatus is located posterior to the male system, with a horseshoe-shaped Lang’s vesicle ( Fig. 8 View FIGURE 8 A). The female gonopore narrows before opening into a common genital atrium, from here the female apparatus extends posteriorly in the form of the vagina externa, which ascends dorsally, while becomes wider before it turns to the vagina media, which is surrounded by abundant eosinophilic cement glands. The vagina media ascends in the form of a dorso-anteriorly running curve to form vagina interna ( Figs. 10 View FIGURE 10 A, B & 11D). The vagina interna after a short distance, with a gentle descending slope ventrally receives the common oviduct ( Figs. 10 View FIGURE 10 A, 11D).
Regrettably, in the holotype specimen, posterior parts of the female reproductive system include the Lang's vesicle and its duct was damaged during sectioning. Therefore observations of these parts are based on the paratype specimens: The vagina interna, then continues as the duct of Lang’s vesicle which descends ventrally, then enters directly into Lang's vesicle. Lang's vesicle has two anterior and one posterior horn, 1.7mm from the common gonopore, is surrounded by abundant cement glands. The inner wall of the vagina is lined with tall and narrow ciliated epithelial cells undrlain by muscle fibers. Cuboidal epithelium supported by muscle fibers covers the duct of Lang’s vesicle. The Lang's vesicle surrounded by a thin layer of muscle fibers ( Fig. 10 View FIGURE 10 A). In the ventral view of paratype cleared specimen, radiating cement glands and the horseshoe-shaped Lang's vesicle are visible behind the common gonopore ( Fig. 7 View FIGURE 7 D).
Differential diagnosis. Based on different characteristics and the character states listed in Table 1 and Figure 12 View FIGURE 12 , the described species differs from all other congeners except D. fulva and D. persica sp. nov. in terms of color patterns. As the species described here lacks distinct color pattern ( Fig. 7 View FIGURE 7 A), while all the other Discocelis species have a color pattern of pale reddish, pale brown, vinaceous cinnamon or fawn background with numerous black or dark brown spots ( Beveridge 2000; Yeri & Kaburaki 1918).
Furthermore, the species presented here is distinguished (details of differences presented in table 1) from D. tigrina in terms of: body shape, distance between the tentacular eye clusters, cerebral eyes patterns, the extent of the marginal eyes, the location of the mouth, the types of prostatoids and the morphology of the vagina. In our second species body is broadly oval, not tapering posteriorly; the tentacular eyes clusters 1.2 mm apart; the cerebral eyes form two clusters each subdivided into anterior and posterior groups; the marginal eyes extend near to posterior end of body; the mouth is located at posterior end of pharynx; two different types of prostatoids exist in terms of size and content; finally the vagina has two dorsal and ventral curves. D. tigrina on the other hand has an oval body tapered posteriorly; the tentacular eyes 2 mm apart; the cerebral eyes form two single elongate clusters; the marginal eyes as a thin band extend up to cerebral organ; the mouth is located in the middle of pharynx; one type, equal in size, of prostatoid exists; and finally vagina has one dorsal and one ventral curve.
The species introduced here differs from D. japonica in terms of: body shape, number of tentacular eyes, distance between tentacular clusters, numbers of post cerebral eyes, extent of the marginal eyes, the location of the mouth, the structure of the male atrium, and the types of prostatoids and the morphology of vagina. In D. japonica : body is tapered posteriorly; tentacular eyes numbered 15–20 in each cluster, 2 mm apart; the posterior cerebral eyes have 15– 16 eyespots; marginal eyes extend up to cerebral organ; the mouth is located in the middle of pharynx; the male atrium has numerous villus-like projections, one type of prostatoid exists; the vagina has one dorsal and ventral curve. Except the similar character states those mentioned above for D. hollemani sp. nov. this new species has: 25–27 tentacular eyes; 8–10 eyespots in its post cerebral group and a male atrium with two distinct lobes.
D. hollemani is different from D. fulva in terms of: cerebral eyes patterns, extent of the marginal eyes, the location of the mouth, the structure of the male atrium, the distribution of prostatoid organs on wall of the male atrium, the types of prostatoids and finally the morphology of the vagina. In D. fulva : cerebral eyes form two single elongate clusters; the marginal eyes extend up to the cerebral organ; the mouth is located in the middle of pharynx; the male atrium has numerous villus-like projections; prostatoids are scattered on the male atrium, posterior lobe's branch of the male atrium, dorsal to vagina, has no prostatoid organs; two different sizes of prostatoids exist; the vagina has one dorsal and ventral curve. In species describe here: cerebral eyes form two clusters each subdivided into anterior and posterior groups; the marginal eyes extend near to the posterior end of the body; the mouth is located at posterior end of the pharynx; two different types of prostatoids in terms of size and content present; prostatoids encircle the posterior lobe of the male atrium; the vagina has two dorsal and ventral curves.
D. hollemani differs from D. pusilla , with regard to: the number of post cerebral eyes (numbered 1–2 in D. pusilla and 8–10 in D. hollemani ); the extent of the marginal eyes (extend to the level of cerebral organs in D. pusilla ), and the structure of the male atrium (the male atrium has numerous villus- like projections in D. pusilla ), the distribution of prostatoids on the male atrium wall (there is few prostatoids in D. pusilla , while prostatoids are numerous in our species); and the morphology of the vagina (with one dorsal and ventral curve in D. Pusilla ).
The species described here also differs from D. parvimaculata in: number of cerebral eyes in anterior and posterior groups (anterior groups have 41–65 eyespots, posterior groups have 5–20 eyespots for D. parvimaculata against 20–25 anterior eyespots and 8–10 posterior eyespots for D. hollemani ); the shape of the male atrium (in ventral view: three distinct lobes exist in D. parvimaculata , while there are two lobes in the species described here); the structure of the male atrium (in sagittal views, there are four distinct lobes in D. parvimaculata , while D. hollemani has two lobes in the male atrium); the distribution of prostatoids on the anterior lob of the male atrium (the tip of the anterior lobe has large prostatoid organs on dorso-ventral sides in D. hollemani , while the anterior tip of the male atrium in D. parvimaculata has not prostatoids on ventral side; and the morphology of the vagina which has one dorsal and ventral curve in D. parvimaculata and two dorsal and ventral curves in D. hollemani .
Finally, D. australis which its taxonomic position will be discussed later with: muscular seminal vesicle, cerebral eyes form single elongate groups, marginal eyes extend to level of cerebral organ and one type of prostatoids is completely different with the species described here.
The two species described here differ from each other in terms of: body shape (the body tapered posteriorly in D. persica , while the posterior end of the body in D. hollemani does not tapered); number of cerebral eyes both in posterior and anterior groups (anterior cerebral groups have 50–52 eyespots and posterior cerebral groups have 25– 30 eyespots in D. persica against 20–25 anterior cerebral eyespots and 8–10 posterior cerebral eyespots in D. hollemani ); the extent of the marginal eyes (marginal eyes extend to the level of cerebral organ in D. persica ; while extend near to the posterior end of body in D. hollemani ); the structure of the male atrium (the male atrium has four lobes in D. persica and two lobes in D. hollemani ); the distribution of prostatoids on the posterior lobe of the male atrium (prostatoids arranged centrally on posterior lobe in D. persica , while encircle the posterior lobe in D. hollemani ); the types of prostatoids (two different sizes in D. persica , two types of prostatoids in terms of size and content in D. hollemani ). Although the spermiducal vesicle was not observed in D. hollemani , but cannot be considered characteristic, because in every specimen the vasa deferentia usually dilate more or less if the animal is sexually mature ( Faubel, 1983; p. 27)
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |