
Micromoina arboricola, Dumont, Henri J., Rietzler, Arnola C. & Kalapothakis, Evanguedes, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3652.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:621598B0-13CE-43A0-AB83-834B87AC6A1D |
|
DOI |
https://doi.org/10.5281/zenodo.5682488 |
|
persistent identifier |
https://treatment.plazi.org/id/03A38785-FFD3-C87B-FF19-AC54FBA0FA58 |
|
treatment provided by |
Plazi |
|
scientific name |
Micromoina arboricola |
| status |
sp. nov. |
Micromoina arboricola sp. nov.
Etymology. the genus name is composed of Moina , with the prefix micro to indicate one of the peculiarities of the new taxon, viz. its small size. The species name is referring to the habitat, tree (holes).
Type locality. A forest in the Medio Rio Doce Valley, Minas Gerais, Brazil (19º45’53,2’’S; 42º37’22,6’’W). Date of collection: March 24, 2012.
Type material. Holotype. A parthenogenetic female on a slide, partly dissected, deposited at the taxonomic collection of the Institute of Biological Sciences at Federal University of Minas Gerais (accession number LTBI 6271).
Allotype. A partly dissected male on a slide, deposited with the holotype (LTBI 6272).
Paratypes. A tube with glycerol, with a mixture of parthenogenetic and sexual females, and few males, in all about 50 specimens, deposited with the holotype (LTBI 6273). Additional paratypes: about ten slides with dissected males and females, provisionally kept in the collection of HJD.
Diagnosis. Extremely small (largest specimens ca 0.56 mm in size). Female antennula long and slender, with sensory seta inserted near base. Male antennule with two–three strongly swollen estaetascs, the remainder small; a strong sensory seta, and a smallest additional, lateral seta. Four endital setae of trunk limb 2 rather short. Postabdomen swollen in both sexes, mostly not constricted at end, without basal spine and with only three feathered setae.
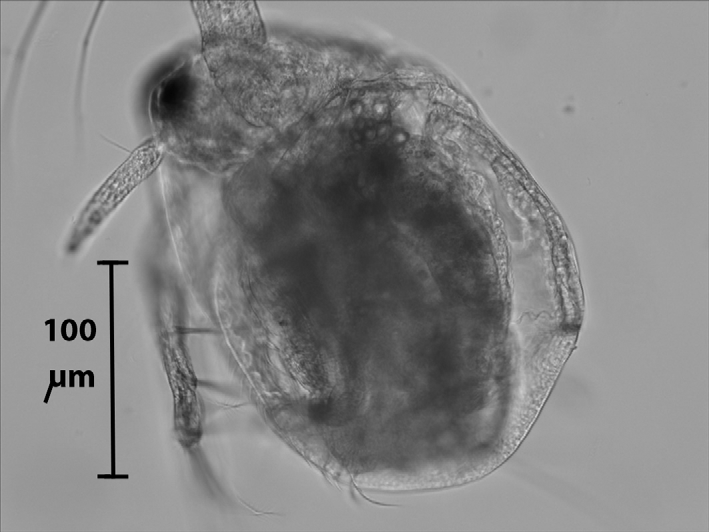
Description. Parthenogenetic female. Body oval and rounded ( Fig. 3 View FIGURE 3 ). Head small, no postocular depression, eyes of moderate size, no ocellus, no hairs on head or valves.
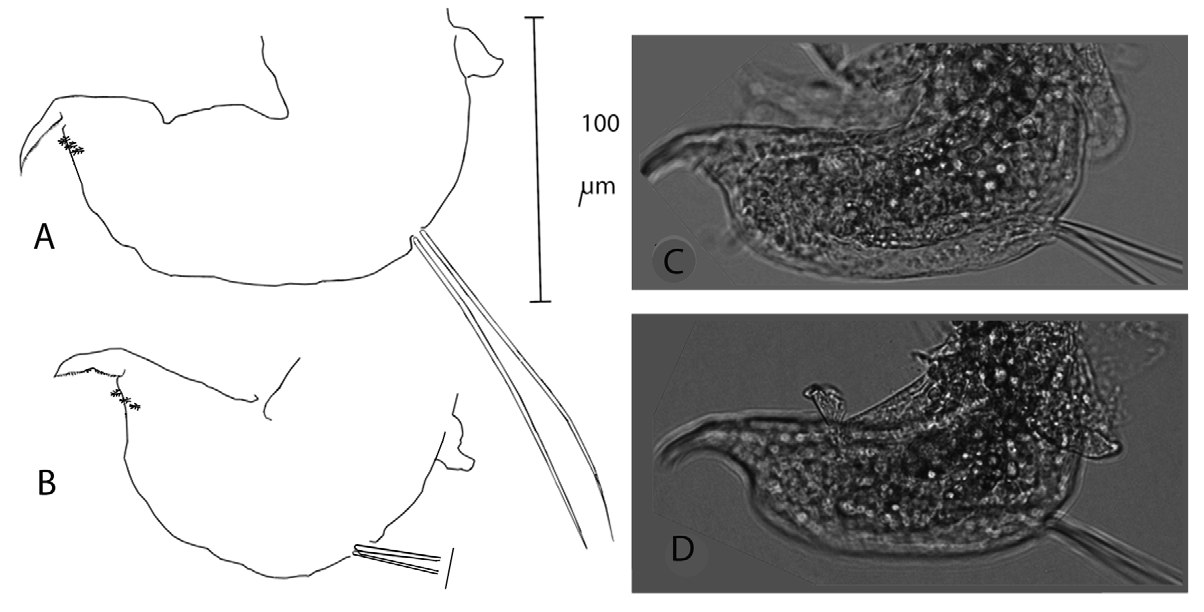
Antennule long and slender, with sensory seta implanted nearer to base than to mid-length ( Figs 3 View FIGURE 3 , 4); nine apical esthaetascs.
Second antenna as for other moinids. The two setae on the apex of the basipodite, between the exo- and endopodite unusually long, the ‘long’ one reaching well over the top of the second endopodite segment, the short one almost reaching the top of the basal segment ( Fig. 7 View FIGURE 7 D). Setae on endopodite with setulation consisting of shortened, almost spiniform setules; setae on exopodite with long and spaced setules.
Labrum with basal segment not swollen, and tip of apical segment narrowed and upturned (of daphniid-type, in Hudec’s terminology) ( Fig. 5 View FIGURE 5 A).
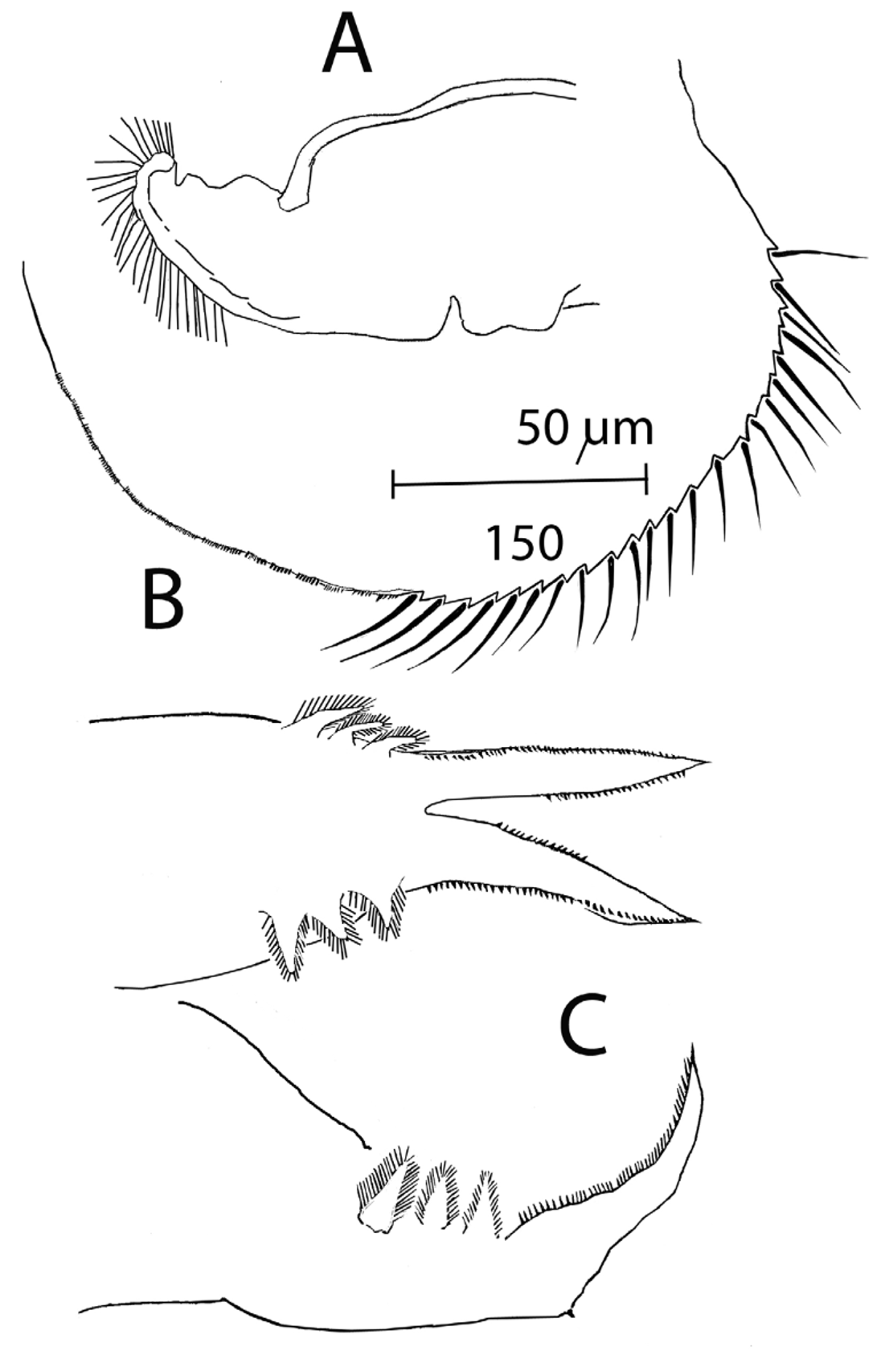
Anterior half of valve rim adorned with a suite of 20-25 rather long, stiff setae, posteriorly replaced by groups of spinules that progressively become smaller and smaller ( Fig. 5 View FIGURE 5 B). Hook-like setae at end of free margin of valves simple, upturned.
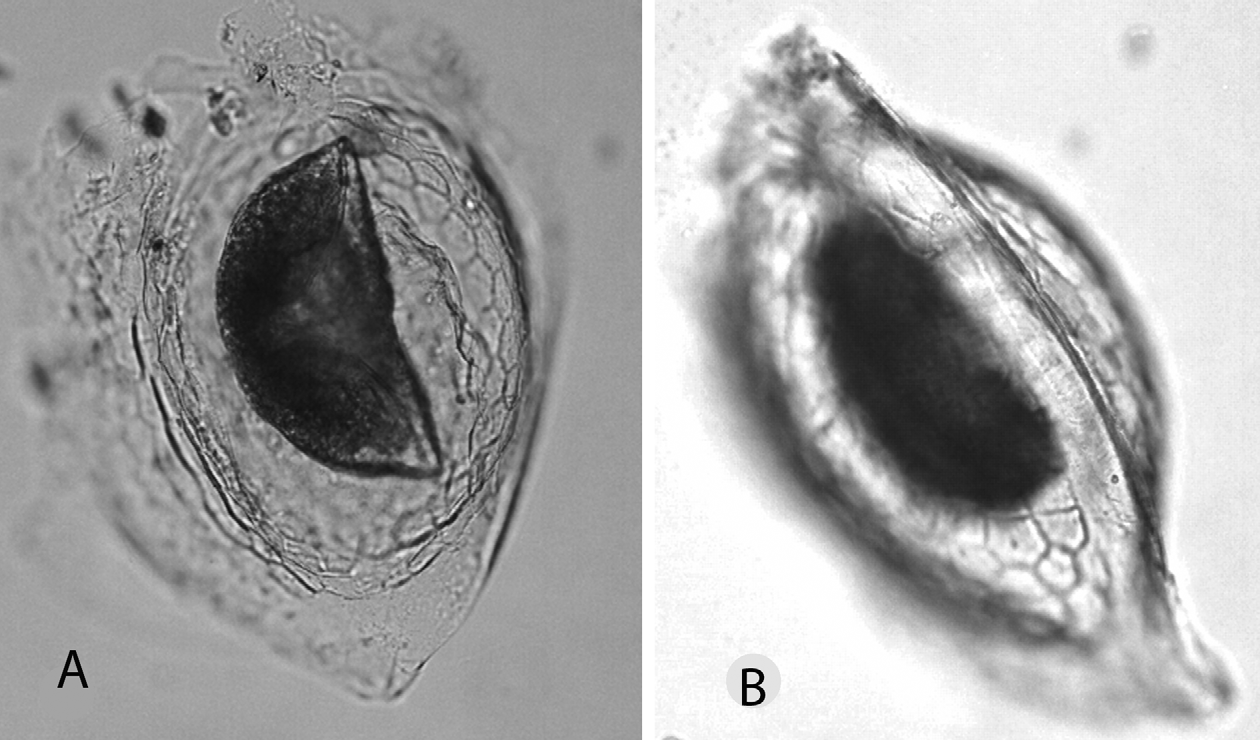
Postabdomen short and plump, relatively variable in shape but lacking the distinctively constricted terminal part typical of all moinids ( Figs 6 View FIGURE 6 ), being in fact more reminiscent of a Megafenestra postabdomen. Lateral appendages reduced to three feathered setae, no basal spine. End-claws short and rather plump, with simple inner and outer pectens, not differentiated into zones.
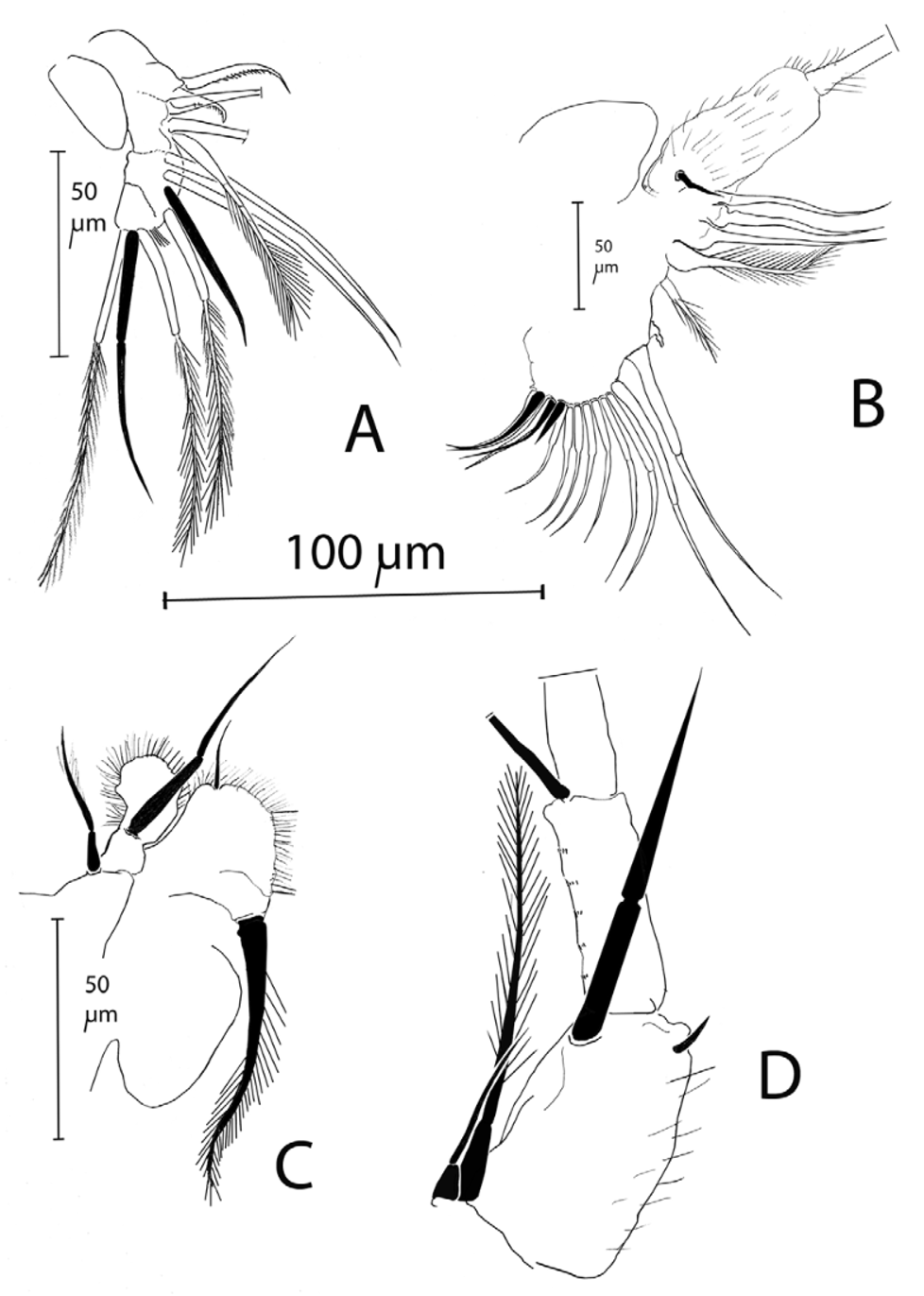
Trunk limbs typical of moinids in general appearance. Trunk limb 1 (T1) almost fully fused, with ten setae of which two ( Fig. 7 View FIGURE 7 A) somewhat shortened and thickened. Ejector hooks well developed, one about twice as long as the other. T2 again typical of moinids (exopodite with one apical seta), but with the four long endital setae not much longer than the basal segment of the exopodite. A short setule at the base of the exopodite. Counting all setae proximad of the small halfway, nipple-shaped appendix on the fused intermediate zone of TL2 as part of the gnathobase (at least functionally), there are about 11 setae in the gnathobasic filter screen, plus three stiff setae in the posterior inner row ( Fig. 7 View FIGURE 7 B). T3 and T4 are as for family. T5 is as for family, with the presumed exopodite bearing two setae (one long, one short), and the inner portion (the ‘endopodite’) reduced to a small lobe, with a seta implanted at half its length. The presumed gnathobase, finally, is another small lobe, bearing one seta as well ( Fig. 7 View FIGURE 7 C).
Ephippium ( Fig. 8 View FIGURE 8 ) rather loosely structured and primitive, containing one egg. Dorsal edge rather sharp. Surface ornamented in pentagons.
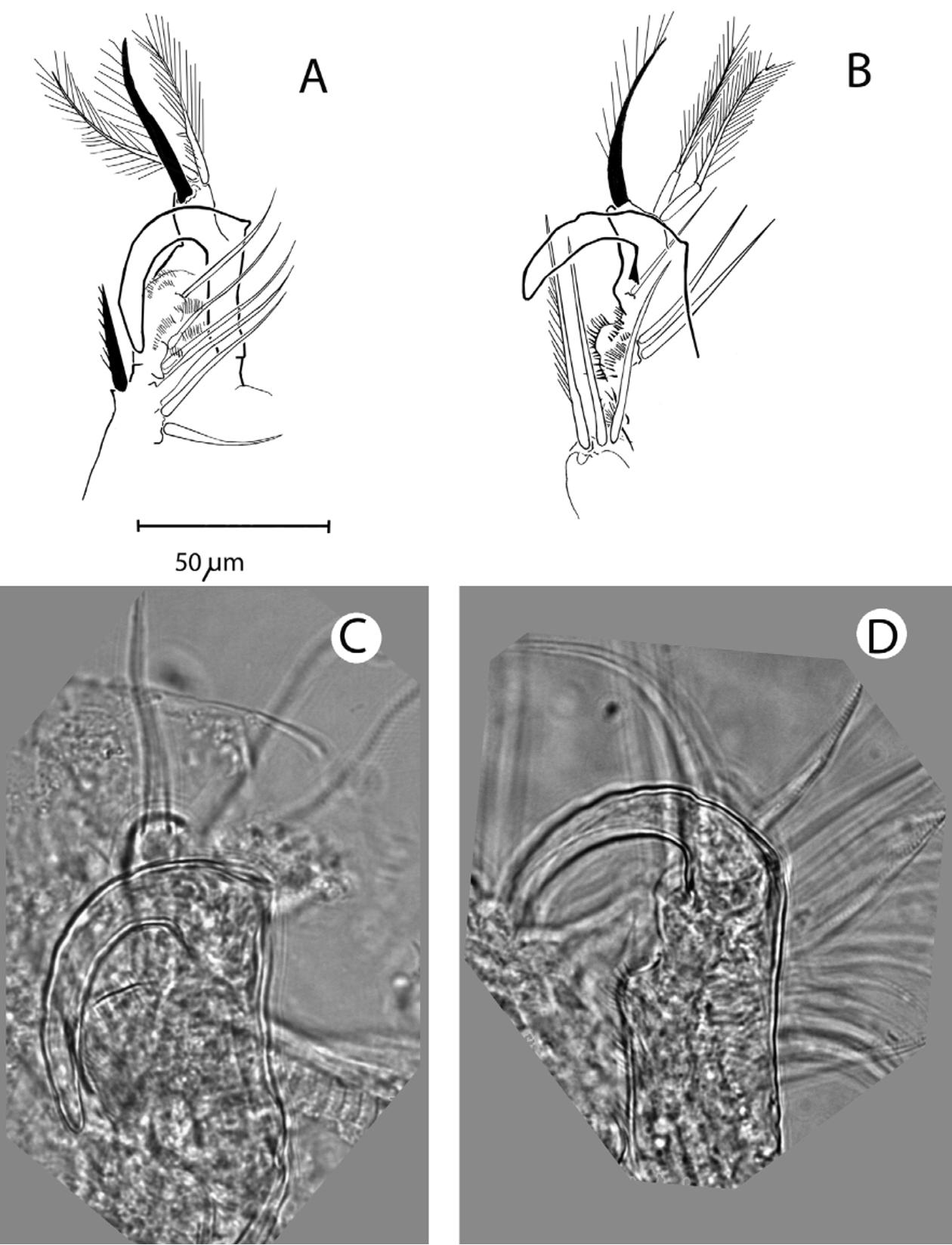
Adult male. Antennule elongated, sharply bent at about 1/3 its length, and provided here with one strong lateral seta and a short and finest extra seta. Apex with hollow esthetascs, of which 2-3 much larger than the others, in all (probably) nine in number ( Fig. 9).
First trunk limb without exopodite; apical segment with three (two slender, one shorter and thicker) plumose setae. A thick copulatory claw with undulate margins ( Fig. 10 View FIGURE 10 ), curved backwards over a somewhat thickened (but variable with angle of view), spinulated fused corm adorned with another six plumose setae, and one stiff seta opposed to the tip of the claw. This entire structure similar to that in all other Moinids with an T1 without exopodite, as most other characters of the male, except its small size.
Size. The smallest moinid known to date. ( Moina minuta , with a total size 0.5-0.7 mm, comes closest). Female: mean body size of 20 specimens from tip of head to posterior tip of valves (0.49 ± 0.04 mm in specimens collected from nature, and 0.55 ± 0.03 mm in cultures set up from such specimens and maintained for 16 weeks (ca 20 generations at 25o C). The difference between both groups is highly statistically different (t-test, P<0.01), i.e. the animals grew larger under optimum food conditions than in nature. Male smaller than female (‘wild’ specimens 0.39 ± 0.04 mm; n=10; lab-reared specimens 0.45 ± 0.05 mm; n=12, the difference between both means statistically significant at P<0.01) but similar in external structure (except antennule) and trunk limbs (except T1).
Molecular information. To situate the new taxon in the phylogenetic tree of its family, we selected several moinids (Exomoina macrocopa (Straus, 1820) , Moina micrura Kurz, 1874 , ‘Moina’ sp. from India, and Moinodaphnia macleayi (King, 1853)) as well as an outgroup composed of three genera of daphniids. We here show two phylogenetic trees, estimated by using a Bayesian approach, and another using the Neighbor Joining technique. Both trees are largely congruent. Relationships between moinid genus-level groups are not well resolved, but the groups themselves are clearly separated. We recover Exomoina, Moina , Moinodaphnia (plus the still undescribed Indian taxon), and Micromoina gen nov. as monophyletic groups within Moinidae , well distinct from Daphniidae ( Figs 11, 12). The polytomy between genera may reflect a breakdown of the phylogenetic signal by saturation, although the genetic distance between them ( Table 1 View TABLE 1 ) does not suggest this.
Exomoina macrocopa Exomoina macrocopa 2 Moinodaphnia macleayi ( Mexico) “ Moina ” sp. ( India)
100.0
63.9
74.9
Micromoina arboricola Moina micrura Daphnia magna Simocephalus vetulus Scapholeberis mucronata Exomoina macrocopa Exomoina macrocopa 2 Moinodaphnia macleayi ( Mexico) “ Moina ” sp. ( India)
0
52.6
1.00
0.78
0.99
Micromoina arboricola Moina micrura Daphnia magna Scapholeberis mucronata Simocephalus vetulus
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |