
Nanotyrannus, Bakker, R. T., Williams, M. & Currie, P. J., 1988
|
publication ID |
https://doi.org/ 10.5281/zenodo.1037529 |
|
DOI |
https://doi.org/10.5281/zenodo.3483102 |
|
persistent identifier |
https://treatment.plazi.org/id/03A1879D-FFD0-FFE9-5E7F-F664FEA6A618 |
|
treatment provided by |
Jeremy |
|
scientific name |
Nanotyrannus |
| status |
gen. nov. |
Nanotyrannus , genus novum
Type species — Gorgosaurus lancensis Gilmore, 1946 View Cited Treatment ( Albertosaurus lancensis View Cited Treatment in Russell 1970).
Type specimen — CMNH 7541 , complete skull and lower jaws in occlusion. Found by David Dunkle in 1942, in the Hell Creek Formation, SE 1/4, sec. 11, R61E, T4S, Sand Creek, Carter County, Montana; Lancian Faunal Age.
Referred specimen — DMNH uncatalogued, three teeth broken off at the crown-root junction, found by D. Reinheimer in 1924, in the Lancian beds of Corson County, South Dakota, accompanied by five juvenile Tyrannosaurus teeth, also broken off at the crown-root junction.
Diagnosis ( Fig. 2 View Figure 2 ). Differs from all other tyrannosaurids in having the following derived features: muzzle width greatly constricted to only one fourth the width of the temporal region; extraordinarily wide basicranium between basitubera and basipterygoid processes; two large pneumatic foramina, placed one behind the other, near the midline of the basisphenoid. Differs from Daspletosaurus , Gorgosaurus and Alioramus and agrees with Tyrannosaurus, Albertosaurus and Tarbosaurus spp. in having the derived features of a marked expansion of the width across the temple, relative to the skull length, and basitubera that are displaced forward towards the basipterygoid processes. Differs from all other tyrannosaurids, except Alioramus , in retaining the primitive characters of a long, low snout and maxillary teeth that are strongly compressed side-to-side. Differs from all other tyrannosaurids in lacking strong ridges and striae along the dorsal surface of the nasal. Differs from Tyrannosaurus in retaining the primitive character of a smooth depressed area for muscle origin on the anterior-ventral corner of the lachrimal.
Etymology» Nano — Latin “small”, plus tyrannus, Latin “tyrant”.
CHARACTER ANALYSIS
In North America there are three successive Land Faunal Ages that have produced tyrannosaurs: the Judithan, the Horseshoe Canyon Faunal Age, and the Lancian. The Lancian is the last fauna of the Cretaceous. Faunal turnover was rapid, and so each age has its diagnostic fauna of common horned dinosaurs and duckbills. The Judithan Age is characterized by the ceratopsians Chasmosaurus and Centrosaurus and by the duckbills Corythosaurus , Lambeosaurus and Prosaurolophus. The Horseshoe Canyon Age is characterized by the ceratopsian Anch- ceratops and the hadrosaurs Saurolophus and Hypacrosaurus. The Lancian is defined by the horned dinosaur Triceratops and the duckbill Edmontosaurus annectens. It would be surprising if large predators had a slower rate of evolutionary turnover, and yet Gilmore’s identification of CMNH 7541 as a Gorgosaurus would give this genus a time range spanning all three faunal ages. No other genus of large dinosaur lasts from the Judithan through the Lancian.
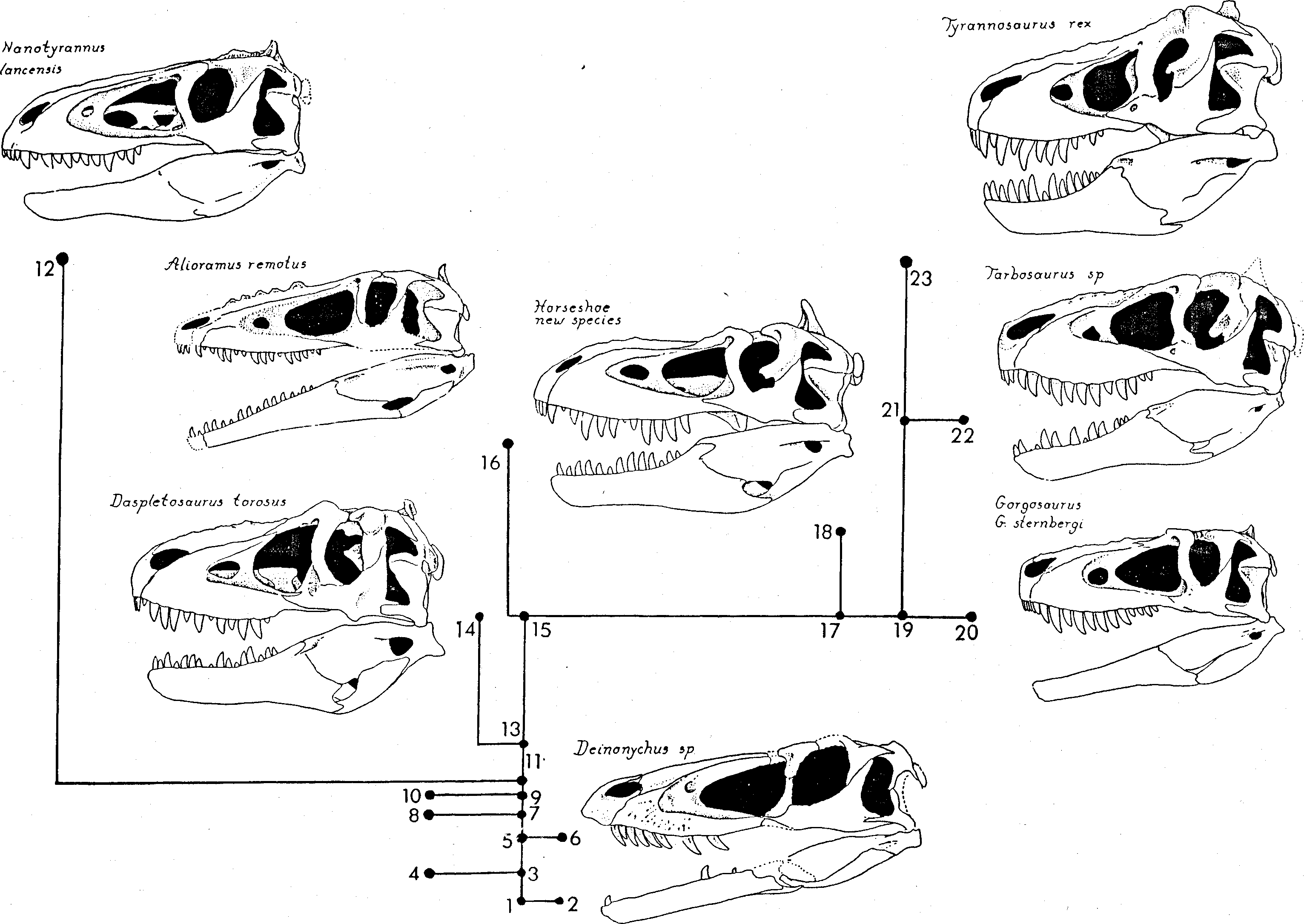
The most recent review of the tyrannosaurids in North America is that by Russell (1970), who recognizes two primitive genera, the gracile Albertosaurus and more robust Daspletosaurus , and one advanced genus, Tyrannosaurus . Both primitive genera, according to Russell, occur in the Judithan Age. Judithan tyrannosaurids are well documented by large samples from the Judith River Formation of Alberta. According to Russell’s review, all named species of Gorgosaurus Lambe 1916 belong within Albertosaurus Osborn 1905 ; Albertosaurus and Daspletosaurus supposedly persisted into the next youngest sample, that of the Horseshoe Canyon. Finally, in the latest sample, from the Lancian Age, Russell identified two genera — a small species which he called Albertosaurus lancensis Gilmore 1946 , and the very large Tyrannosaurus rex, Osborn 1903 . Russell thus gave the genus Albertosaurus the time range of Judithan through Lancian, and this anomalously great generic longevity remained unexplained. The Mongolian Late Cretaceous Nemegt Formation has yielded two types of tyrannosaurid — Tarbosaurus Maleev 1955 , intermediate in structure between Albertosaurus and Tyrannosaurus , and Alioramus Kurzanov 1976 , a strange genus with a long snout ornamented with curious excrescences.
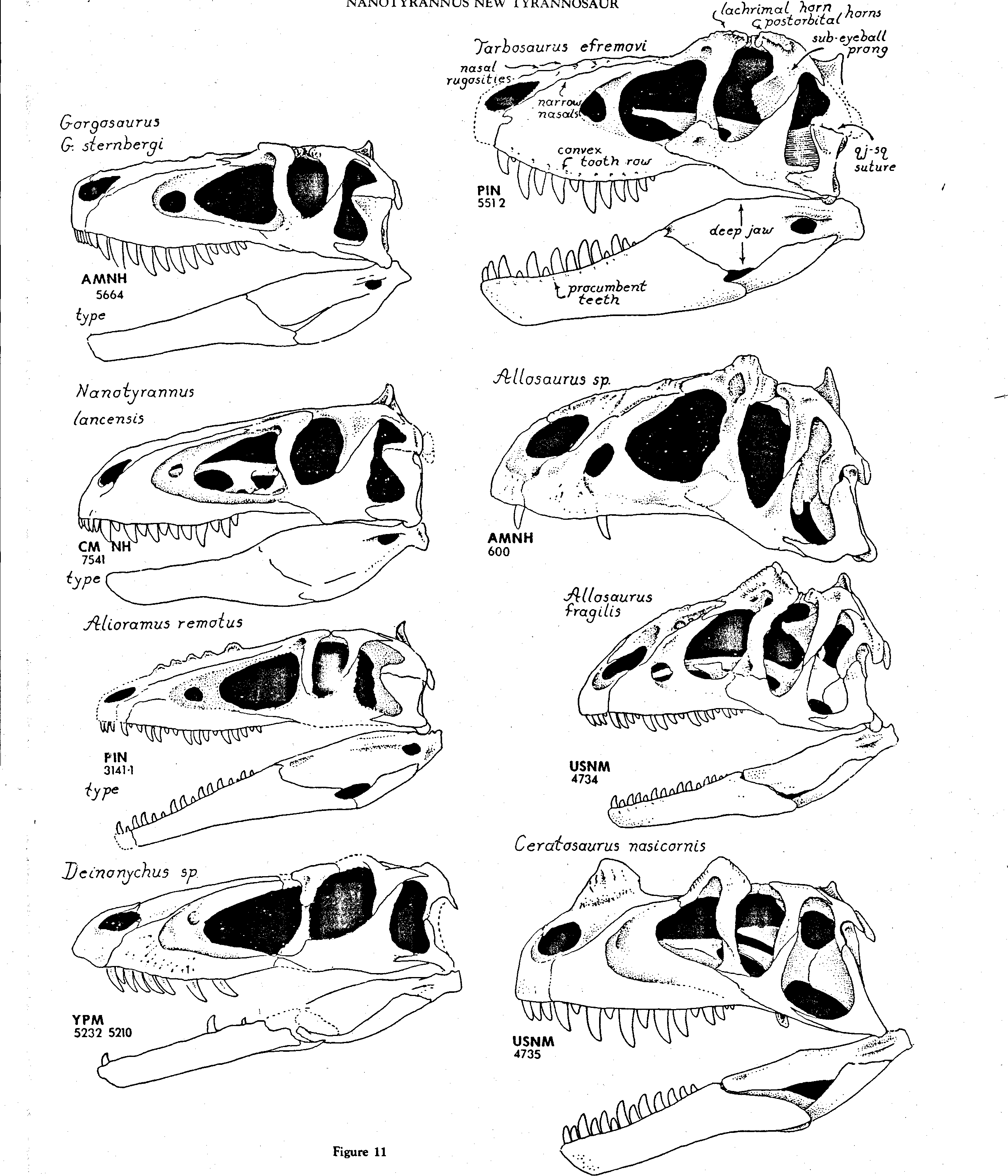
As we shall show in a subsequent publication (Bakker, Currie, and Williams, in press), the genus Albertosaurus is distinct from the genus Gorgosaurus , and these two genera, plus Tarbosaurus and Tyrannosaurus , make a clade distinct from Daspletosaurus . In the following analysis, we restrict the concept of Albertosaurus to the type and paratype specimens NMC 5600 & 5601, both from the Horseshoe Canyon Formation. A great variety of tyrannosaurs have been grouped together in Gorgosaurus and we are confident that at least two species can be diagnosed ( Fig. 11 View Figure 11 ). In the Judith River sample there is a gorgosaur, represented by FMNH 308, with relatively small teeth and a high tooth count (15 in the maxilla). In addition, there is a Judith River gorgosaur, represented by USNM 12814 and AMNH 5336, with relatively large teeth and a low tooth count (13 in the maxilla' Both of these categories have robust cranial bones and occipital condyles that are depressed top-to-bottom, suggesting that move ment of the head on the neck was concentrated in a horizontal plane. Several additional Judithan gorgosaur specimens, such as ROM 1247 and AMNH 5664, have unusually delicate braincases and occipital condyles that are spherical and not depressed. Russell (1970) identified these delicate gorgosaurs as juveniles of the robust species but they may, in fact, represent a distinct species, Gorgosaurus stembergi Matthew and Brown 1922 . Unfortunately the type specimen of Gorgosaurus is the type of G. libratus Lambe 1916, a skeleton that lacks a braincase and is very badlv broken in the muzzle, and the correct nomenclature for gorgosaurs is hard to work out. Therefore, in the following discussion, we will use the informal categories “gracile gorgosaur”, “small-tooth gorgosaur”, and “ big-tooth robust gorgosaur”. A new genus of gorgosaur-like tyrannosaurid is represented by Tyrrell Museum P 81.3, a nearly complete skull and much of the postcranium from the Horseshoe Canyon Formation. This skull shows a mixture of features seen separately in Alioramus , Gorgosaurus , and Tarbosaurus .
.
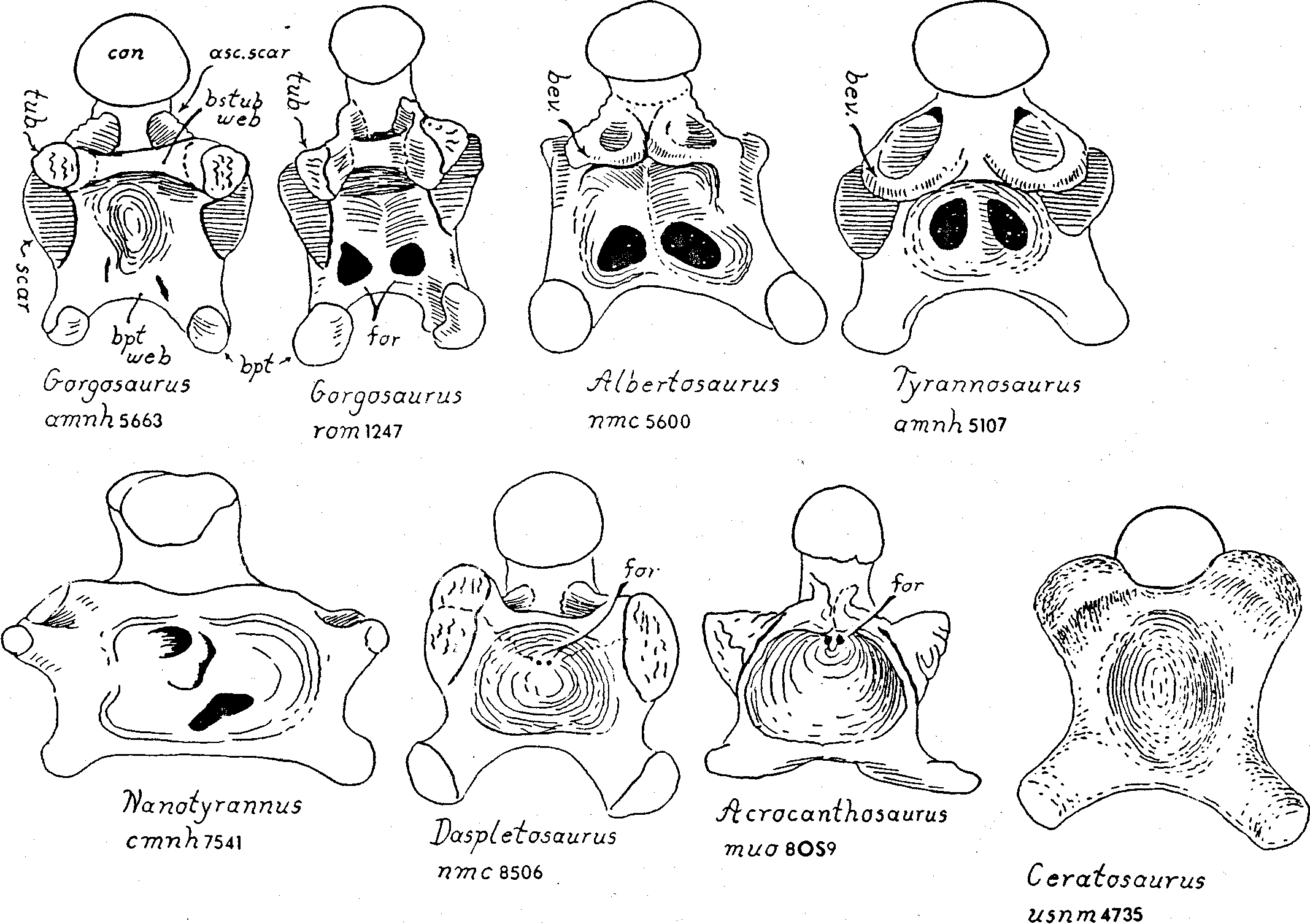
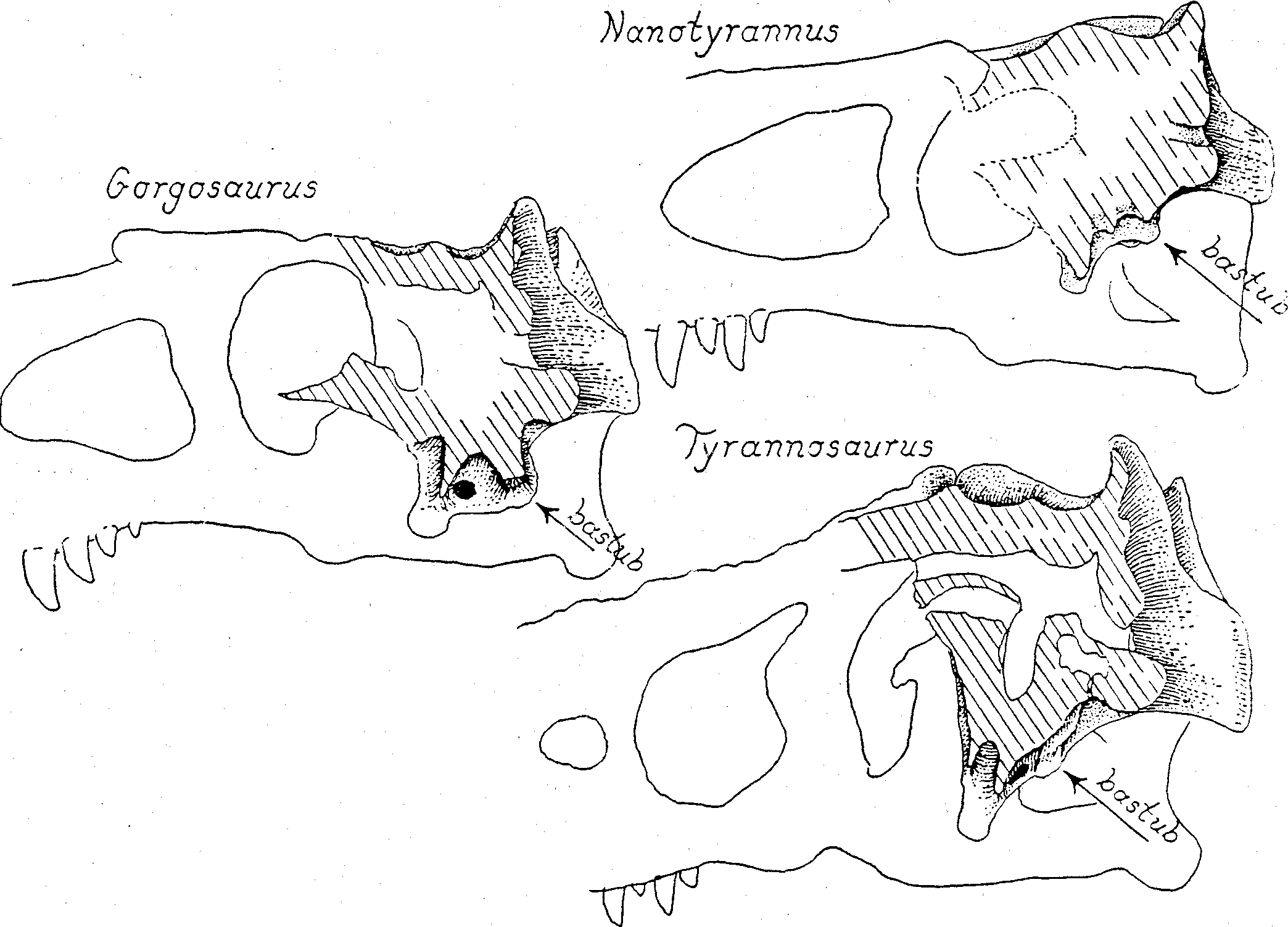
Basicranium. We introduce here a few descriptive terms that are useful in comparing theropod braincase structures. The transverse web of bone connecting right and left basipterygoid processes is the “basipterygoidal web” (bpt.web in Fig. 4 View Figure 4 ). The transverse web of bone connecting right and left basituberal processes is the “basituberal web” (bastub.web in Fig. 4-6 View Figure 4 View Figure 5 View Figure 6 ). The wall of basisphenoidal bone running fore-to-aft connecting the basipterygoid web to the basituberal web on either side is the “basicranial boxwork wall” (boxwork wall in Fig. 5 View Figure 5 ). The two boxwork walls and the two transverse webs constitute the “basicranial boxwork”.
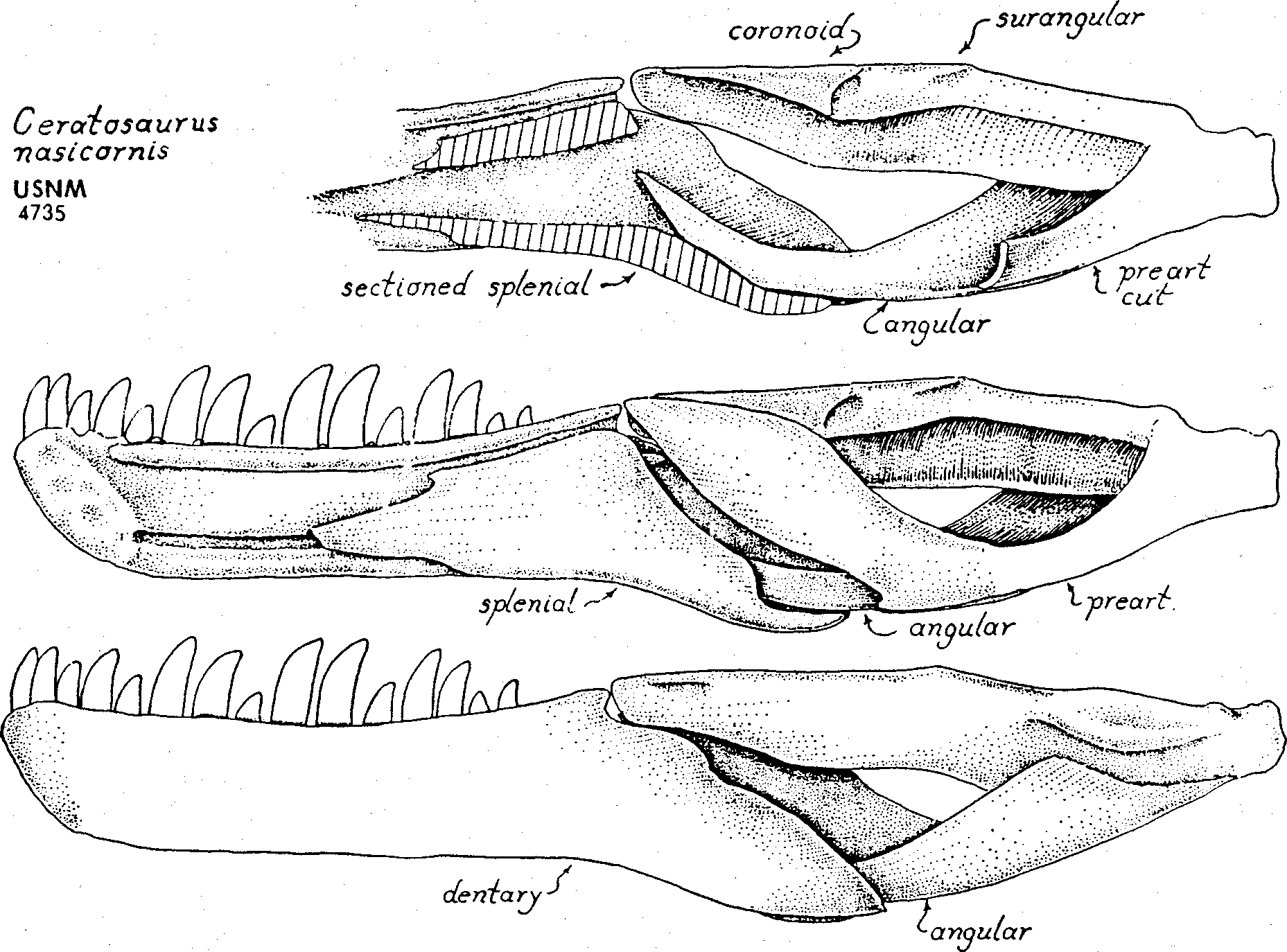
The most outstanding feature of Nanotyrannus is the enormous lateral expansions of the basicranial boxwork, relative to the length of the skull ( Fig. 3-5 View Figure 3 View Figure 4 View Figure 5 ). In primitive theropods, such as Ceratosaurus , the width across the basitubera is only about 8% of the skull length (USNM 4735; Fig. 4; Table 1). In daspletosaurs and in most gorgosaurs the basituberal width reaches 12% to nearly 15% of the skull length. In Tyranno * saurus the basituberal width is enormously expanded, reaching 19% of the skull length ( Fig. 4-6 View Figure 4 View Figure 5 View Figure 6 ; Table 1). In Nanotyrannus the basituberal width is fully 20% of skull length as we restore the skull, and the condition of the specimen strongly suggests that the basicranium has been preserved with no distortion of width. Such lateral expansion of the basitubera as is seen in Nanotyrannus would greatly increase the leverage available to the neck muscles for flexing the head sideways.
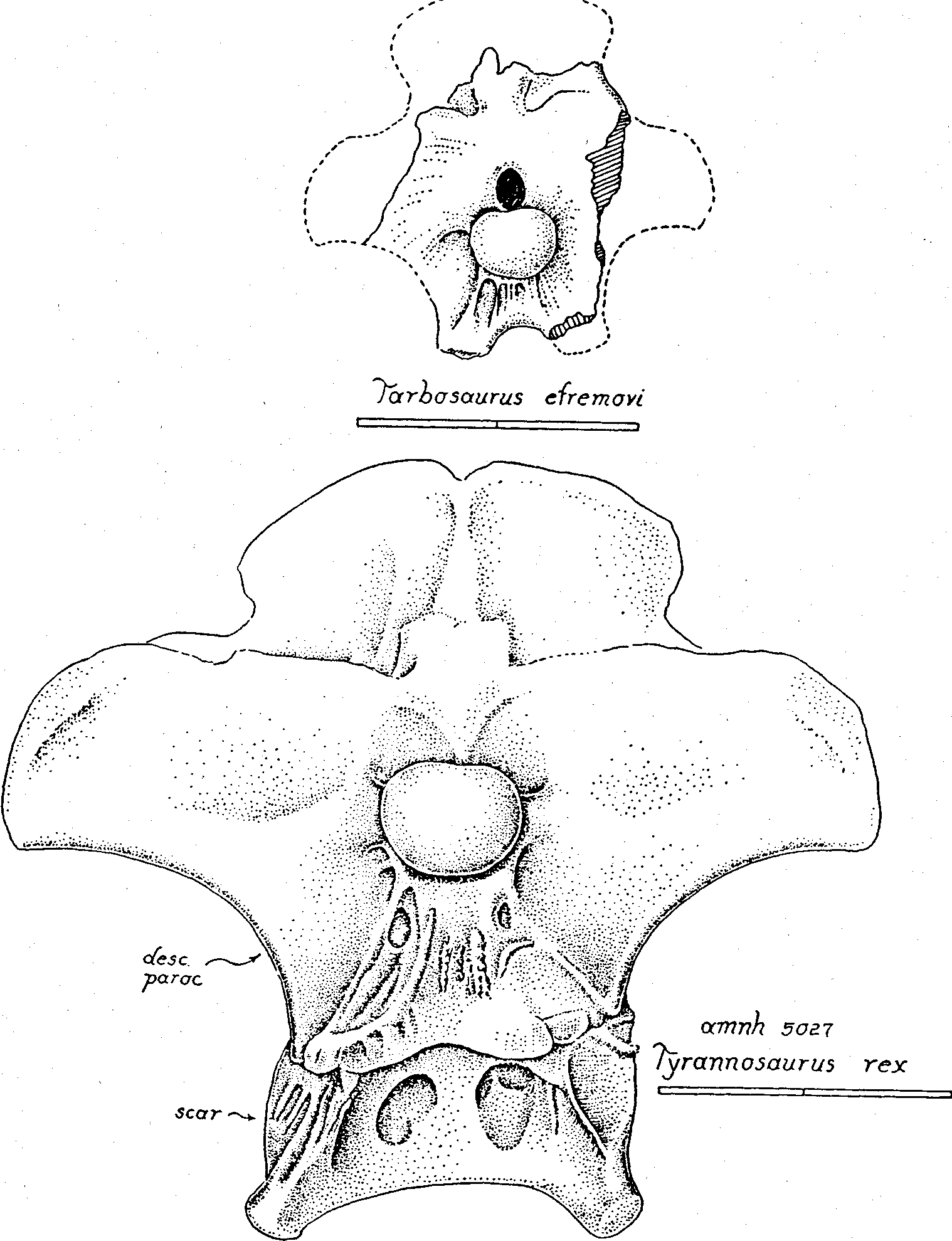
The basitubera in tyrannosaurs are composed of three units: 1) The main tuberous process (tub. in Fig. 4-5 View Figure 4 View Figure 5 ) is a swollen development of the basioccipital at the posterior-outer corner of the basicranial boxwork. In gorgosaurs and daspletosaurs the tuberous process is very thick but in Albertosaurus , Tyrannosaurus and Nanotyrannus it is reduced in bulk, a change that reflects a decrease in the size of the muscle tendon attaching to the process. 2) A line of rugose muscle attachment scars ascends the posterior face of the basioccipital from the tuberous process. This ascending scar complex is well developed in Nanotyrannus , as it is in most other tyrannosaurs, and marks the attachment of the longus colli muscle (= rectus capitis anterior of human anatomy). 3) Anterior to the tuberous process, along the ventral-lateral rim of the basicranium, is a large, flat, oval scar (scar in Fig. 4-6 View Figure 4 View Figure 5 View Figure 6 ) marking the insertion of the iliocostalis cervicis capitis muscle. This oval scar is small and weakly striated in Nanotyrannus , compared to the state in other tyrannosaurs, suggesting that the muscle attachment was fleshy rather than tendinous.
In primitive theropods and most tyrannosaurs, the ventral surface of the basisphenoid is excavated by a huge central cavity (cav. in Fig. 5 View Figure 5 ) bounded by the basipterygoidal web in front, the basituberal web behind, and the basicranial boxwork walls on either side. Foramina of various sorts penetrate the ceiling of this cavity. In primitive theropods the cavity is very deep and a pair of tiny vascular foramina penetrate the anterior part of the cavity, just anterior to the basituberal web, a condition retained in allosaurids, dromaeosaurids and acrocanthosaurs ( Fig. 4, 9 View Figure 9 ). Among tyrannosaurids this primitive state is known only in Daspletosaurus ( Fig. 8 View Figure 8 ). In the long-snouted Mongolian tyrannosaurid Alioramus and in the new genus of tyrannosaurid from the Horseshoe Canyon the paired foramina are displaced forward but are still relatively small. In gorgosaurs the foramina are anterior in position and greatly enlarged. These enlarged foramina lead to a complex sytstem of pneumatic spaces within the braincase. In Albertosaurus and Tyrannosaurus the foramina are still larger than those in gorgosaurs ( Fig. 4-6 View Figure 4 View Figure 5 View Figure 6 , 11 View Figure 11 ). Nanotyrannus has a unique foraminal pattern — there is one large foramen anteriorly almost on the ventral midline of the basisphenoid and a second large foramen, with an irregular outline, off-center further posteriorly in front of the basituberal web ( Fig. 3 View Figure 3 , 4). Preparation of the matrix filling the foramina is not yet complete, but one or both almost certainly lead inward to the pneumatic spaces. It is not obvious how the Nanotyrannus system could evolve from the gorgosaur pattern of bilaterally symmetrical and paired foramina. It is quite possible that the pneumatic system of Nanotyrannus evolved independently from the gorgosauralbertosaur-Tyrannosaurus clade.
Most Late Cretaceous theropod dinosaur families show trends for increasingly complex pneumatization of the braincase ( Currie, 1985; Bakker, Currie, and Williams, in press). The functional significance of large pneumatopores is not simply to “lighten the skull” because hollow bones have evolved in some birds without the development of large pneumatic foramina. Furthermore, all modern crocodilians have skulls full of pneumatic spaces and yet the pneumatic foramina opening to the outer skull bones are all very small. Clearly large pneumatopores are not necessary for the development of thin-walled, hollow bones. The relatively huge foramina in Nanotyrannus , Gorgosaurus , Albertosaurus and Tyrannosaurus suggest that there was easy movement of air through the holes. Some modern predators, such as the cheetah (Acinonyx jubatus), incur great exercise heat loads in their heads during running or subduing struggling prey (Taylor et al., 1973). A relatively small brain, such as that possessed by tyrannosaurs, could reach lethal temperature more rapidly than a large brain. It may be the air from the throat was circulated up into the basicranium in tyrannosaurs, and the air was cooled before entry by evaporation along the walls of the mouth. Alternatively, the large foramina may be part of adaptations for increasing the sensitivity of the ears to certain frequencies ( Currie, 1985). These explanations are not mutually exclusive, and both cooling and auditory functions may be part of the adaptive design in tyrannosaurs.
In primitive theropods and in gorgosaurs the ceiling of the central basisphenoidal cavity is very tall and the cavity is very deep top-to-bottom. The cavity remains very deep in daspletosaurs and gorgosaurs, and in gorgosaurs the large pneumatic foramina face mostly inwards from their location on the steep inner surfaces of the boxwork walls ( Fig. 4-6 View Figure 4 View Figure 5 View Figure 6 , 8 View Figure 8 ).
In Nanotyrannus the ventral floor of the basisphenoid is nearly flat between the basituberal and basipterygoidal webs, a condition seen in Tyrannosaurus and, to a lesser extent, in Albertosaurus . In the last two genera and, probably, in Nanotyrannus as well there are very large pneumatic chambers above the basisphenoidal floor, filling in the space that is occupied by the primitive open cavity in gorgosaurs ( Fig. 3-6 View Figure 3 View Figure 4 View Figure 5 View Figure 6 , 9 View Figure 9 ).
The occipital condyle is carried on a constricted neck in Nanotyrannus and the neck faces far more strongly downward than in gorgosaurs and daspletosaurs, or in ceratosaurs and allosaurs, agreeing with the condition in Tyrannosaurus . Moreover, the articular surface on the con dyle in Nanotyrannus has been displaced downwards and around onto the ventral surface of the neck far more than in gorgosaurs and daspletosaurs, again agreeing with Tyrannosaurus ( Fig. 6 View Figure 6 , 9 View Figure 9 ). Clearly the head was flexed far more sharply downwards in Nanotyrannus and Tyrannosaurus than in more primitive tyrannosaurids. The type and paratype specimens of Albertosaurus are badly broken but, as far as is known, Albertosaurus approaches Tyrannosaurus in these adaptations for head-neck flexure and is distinctly more advanced than Gorgosaurus . No complete braincase has been described for the Mongolian Tarbosaurus but the available evidence suggests that the structure is inter mediate between Albertosaurus and Tyrannosaurus ,
The orientation of the basitubera must change to maintain leverage when there is an evolutionary increase in the flexure at the head-neck joint. In an animal that frequently holds its head out straight, in line with the cervical vertebrae, such as a modern crocodilian, the basitubera must project directly downwards to maximize the leverage of the longus colli pull ing directly backwards ( Fig. 7 View Figure 7 , 10 View Figure 10 ). But, in an animal carrying its head flexed at nearly a right angle to the neck, such as an ostrich, the basitubera must be displaced forward along the basicranium to achieve the same leverage. With the head flexed at a right angle to the neck, the basitubera need not project downwards at all, because leverage of the longus colli is controlled entirely by the distance from the center of rotation of the condyle to the floor of the braincase at the base of the basitubera. In primitive tyrannosaurs, such as the gracile gorgosaurs, the basitubera are located far aft, beneath the occipital condyle, and project strongly downward ( Fig. 7 View Figure 7 , 10 View Figure 10 ). But in Tyrannosaurus and Nanotyrannus , the basitubera are displaced forward and project less strongly below the floor of the basicranium. Thus these two genera were equipped for powerful head-neck movements in a vertical plane in an arc that was, on average, far more flexed than in gorgosaurs or in primitive theropods generally.
The occipital condyle, as seen from behind, is triangular, with rounded comers, in Nanotyrannus , and resembles that of gracile gorgosaurs rather than the dorsal-ventrally depressed condyle of Tyrannosaurus rex and some robust gorgosaurs ( Fig. 3 View Figure 3 , 5 View Figure 5 , 6 View Figure 6 , 9 View Figure 9 ). The triangular shape suggests that side-to-side movement was more extensive when the head was extended in line with the neck than when it was sharply flexed downwards.
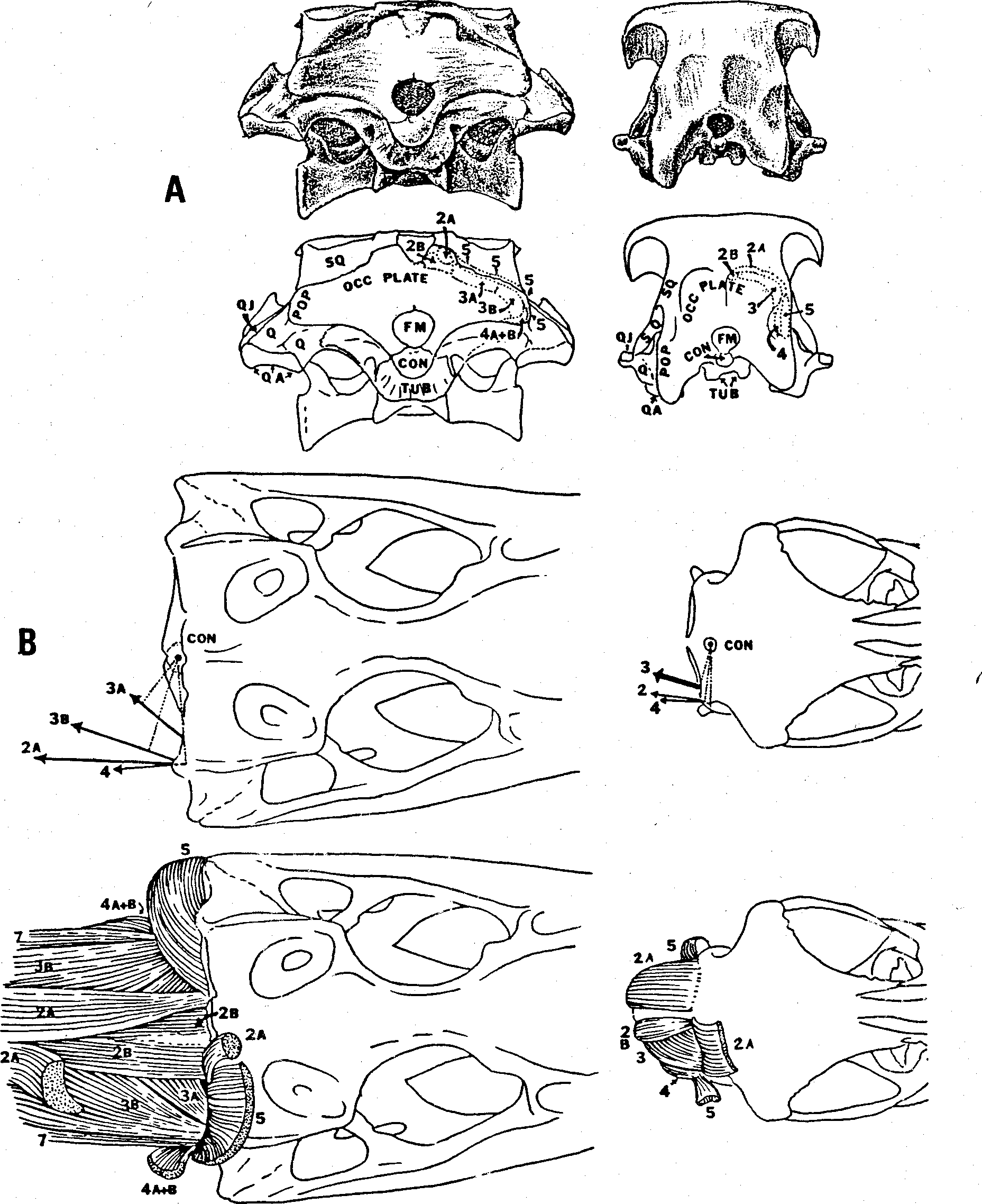
In Nanotyrannus , the occipital plate above the foramen magnum is reinforced by a vertical midline wedge of supraoccipital bone (wedge in Fig. 5 View Figure 5 ) that is capped dorsally by two rounded tab-like processes placed side by. side. The tandemcap arrangement is unique to the Tyrannosauridae and found in all genera of the family ( Fig. 3 View Figure 3 , 5 View Figure 5 , 6 View Figure 6 , 9 View Figure 9 ). The supraoccipital wedge received the insertion of the short rectus capitis (= rectus capitis posterior of human anatomy) running forward from the top of the axis neural spine; the two caps probably received two tendons of that muscle. Above and either side of the tandem caps in Nanotyrannus , as in other tyrannosaurs, is a great expanse of the parietal wings (pa. wing in Fig. 5 View Figure 5 ). The central part of the posterior face of the parietal wing is concave and must have received the insertion of a thick fleshy bundle of the long rectus capitis originating from the tops of the cervical neural spines. The dorsal margin of the parietal wings is thickened and must have received the insertion of the spinalis capitis, the most dorsal cervical muscle. In North American tyrannosaurs other than Nanotyrannus , the outer corners of the parietal wings are rounded in contour and are ornamented with serrated rugosities, indicating a tendinous attachment of the outer part of the spinalis capitis muscle. But in Nanotyrannus the dorsal margin of the parietal wing meets the lateral margin at a sharply defined angle and the rugosities are poorly expressed, indicating a less tendinous attachment here. Right and left parietal wings meet above the supraoccipital wedge at the dorsal occipital midline and the bone here is thickened and striated in a vertical direction, indicating the attachment of a thick tendinous sheet that must have separated the right and left halves of the spinalis capitis-rectus capitis bundles.
.
Temporal Region. In Nanotyrannus the width of the superior temporal fenestrae and of the entire temporal region is greatly expanded relative to the snout ( Fig. 3 View Figure 3 , 11 View Figure 11 ). The breadth between the jaw joints, measured between quadratojugals, is 57% of the skull length; the breadth of the snout at the midpoint of the main antorbital fenestra is only 13% of the skull length, so that the quadratojugal width/snout width ratio is 4.3 ( Table 1). In the best preserved Tyrannosaurus skull, AMNH 5027, the quadratojugal width is 62% of skull length but the muzzle is not nearly as constricted as in Nanotyrannus and the ratio of quadratojugal width to mid-snout width is only about 2. No gorgosaur or daspletosaur skull known shows a ratio of quadratojugal width to skull length or to muzzle width anywhere nearly as high as in Nanotyrannus . The type of Nanotyrannus has been subjected to slight postmortem distortion — the occiput is pushed forward slightly, crushing the quadrate forward, and the snout is twisted about its long axis so that the left external naris is rotated to the right. But in general the specimen shows virtually no compression lines or fractures and the extraordinary constriction of the muzzle cannot be ascribed to postmortem deformation.
The seven best Judithan Age gorgosaur specimens are distorted in various directions but not one even approaches the shape of Nanotyrannus ( Fig. 11 View Figure 11 , Table 1). All the available evidence indicates that dorsal-ventral compression, suffered by a theropod skull during the processes of burial and sediment compaction, does not expand the skull width at the rear. The type of Gorgosaurus stembergi (AMNH 5664) is compressed from top-to-bottom but this distortion has simply collapsed the skull roof down onto the bones along the jaw margin — there has been no spreading of the skull between the jaw joints and the quadratojugal breadth is only 39% of the skull length. In AMNH 5336, a robust, big-toothed gorgosaur, the skull has suffered moderate dorsal-ventral distortion that has compressed the parietal wings of the occiput downward but the quadrates remain nearly vertical and the quadratojugal breadth is only 39% of the skull length. In USNM 12814, a gorgosaur skull preserved with slight oblique distortion in a dorsal-lateral direction, the quadratojugal breadth is 42% of the skull length. This skull is probably the least distorted known for a gorgosaur and shows a sharply defined break in the skull contour, as seen in dorsal view, between the muzzle and the temporal region, but the degree of temporal expansion is very weak compared to that seen in Tyrannosaurus and Nanotyrannus . The type of Daspletosaurus torosus , NMC 8506, is preserved with no visible distortion (Russell, pers. comm. 1988); the muzzle is wider than that of gorgosaurs, and the temporal region is expanded only slightly relative to muzzle breadth. In the gracile gorgosaur skull ROM 1247, moderate side-to-side compression has reduced the quadratojugal breadth to 26% of skull length; in life the ratio was probably about 37%. The robust, smalltoothed gorgosaur skull FMNH 308 was chosen by Russell (1970) to illustrate the structure of common Judithan species G. limbatus, but this specimen is severely distorted side-toside, with the quadratojugal breadth only 25% of the skull length; in life the ratio was probably similar to that of ROM 1247. The robust, big-toothed gorgosaur skull AMNH 5458 (the specimen mounted in a running pose in the public gallery of the American Museum) shows a little evidence of modest lateral distortion and has a quadratojugal width 33% of skull length.
It is especially important to note that, of the seven best Judithan skulls, three show slight to severe side-to-side compression (FMNH 308, AMNH 5434, ROM 1247) and none show strong top-to-bottom compression. In contrast, the sample of Lancian tyrannosaurid skulls shows no example of side-to-Side distortion — the type of Nanotyrannus and the three best Tyrannosaurus skulls, AMNH 5027, LACM 23844, and Tyrrell Museum P 81.6, are undistorted or show slight to very slight dorsal-ventral compression and very slight front-to-back shear. Surely the natural shape of the skull will determine the pattern of postmortem distortion — after death, skulls that are relatively tall and narrow will tend to fall on their sides and be so buried, whereas skulls that are wide will tend to rest right-side-up on the substrate.
In the type of Nanotyrannus the quadrates have a slight outward-and-downward slant as seen in posterior view ( Fig. 3 View Figure 3 ), a condition repeated in Tyrannosaurus and in the best preserved gorgosaurs and daspletosaurs (USNM 12814, NMC 8506). As in all tyrannosaurs, the Nanotyrannus quadrate is very wide distally relative to the shaft length so that distal width is only slightly less than maximum quadrate height. In primitive theropods such as podokesaurs and ceratosaurs the quadrate shaft is much longer relative to the distal articular width. Allosaurs have a quadrate that is very close in proportions to that of tyrannosaurs.
As a consequence of the temporal breadth/muzzle breadth ratio in Nanotyrannus and Tyrannosaurus , right and left eyes have a wide overlap of visual fields. Stereoscopy was probably present over a range of 30 degrees or more. Since the orbits face upwards as well as forwards in Tyrannosaurus , the widest possible arc of visual field overlap lies in a plane that is tilted upwards and forwards relative to the muzzle. In Nanotyrannus the upward orientation of the orbits is less marked, and, because the muzzle is extremely constricted, a wide arc of overlap is possible in a plane almost parallel to the muzzle.
The paroccipital processes of Nanotyrannus agree with those of Tyrannosaurus in being oriented nearly directly outwards and having much less of a backward orientation than that seen in gorgosaurs and daspletosaurs ( Fig. 3 View Figure 3 , 11 View Figure 11 ). The area for muscle attachment in the superior temporal fossa is very great in Nanotyrannus , as it is in all tyrannosaurids: The right and left temporal muscle scars meet along a very high and very narrow sagittal crest that extends from parietal at the front face of the occipital plate forward along the frontal (midline to a point opposite the dorsal midmargin of the orbit. The anterior extension of the temporal scars in tyrannosaurs is developed as an excavated area on the dorsal surface of the frontals and is far larger than that of the large theropods of the Early Cretaceous or Jurassic, such as Acrocanthosaurus, allosaurs and ceratosaurs. The sutures between the lachrimal and prefrontal have thoroughly coalesced in Nanotyrannus , as have the sutures between frontals and prefrontals, and between parietals and frontals. This degree of coossification is not matched in any gorgosaur or daspletosaur specimen and is even greater than in the Tyrannosaurus skull AMNH 5027. Without question, the type of Nanotyrannus was fully adult and had reached the maximum size the individual would have attained if it had lived longer.
The lower temporal fenestra in the type of Nanotyrannus has suffered slight fore-to-aft compression but appears to have been like that of tyrannosaurs in general, with a long, horizontal quadratojugal-squamosal contact that gives a W W-shape to the posterior outline of the fenestra. Although the Nanotyrannus type is clearly fully adult, there is no coossification at the squamosal-quadratojugal contact, and, since the quadrate has a moveable joint with the squamosal, the entire quadrate-quadratojugal complex can swing outwards to increase the size of the gullet. The quadratojugal in Nanotyrannus has a long anterior prong that overlaps onto a deep notch in the posterior process of the jugal. Postmortem compression has pressed the quadratojugal prong forward so that it extends beyond its jugal articular surface. There is no coossification at this joint, suggesting that some twisting and flexing could occur in life, a situation that also occurs in Tyrannosaurus and the other genera of the family. The postorbital bar in Nanotyrannus , as in tyrannosaurids generally, is very wide front-to-back with a firm, interdigitating suture between postorbital and jugal ( Fig. 3 View Figure 3 , 11 View Figure 11 ). The postorbital bar in all Jurassic theropods is much narrower. The postorbital bar in the troödont Saurornithoides is very similar to that of Nanotyrannus and the other tyrannosaurids ( Barsbold, 1974). Although the tyrannosaurid post' orbital bar is very wide, it is thin side-to-side and some bowing or twisting is possible about the postorbitabjugal sutural zone. Below the posterior-ventral edge of the orbit in Nanotyrannus , the jugal bears a Slight swelling with a lightly striated surface, indicating the presence in life of a low dermal horn. A horn in this location is present in allosaurs and most Cretaceous theropods of large size but appears to be absent in ceratosaurs and primitive theropods of the Late Triassic and Early Jurassic, such as Coelophysis and Syntarsus.
Orbit and Muzzle. The lachrimal in Nanotyrannus was restored in plaster with a pre-orbital horn of a most unusual configuration ( Plate 1-3 View Plate 1 View Plate 2 View Plate 3 ) — in side view, the horn has an evenly rounded, convex-up profile. When the plaster was removed, the dorsal surface of the lachrimal was seen to lack any clearcut horn-like growth, although further preparation is necessary to clean down to the bone. At present, it appears that the dorsal preorbital bar is neither greatly swollen nor highly rugose. Ceratosaurs and allosaurs have erect horns with strong rugosities ( Fig. 11 View Figure 11 ). All gorgosaurs and daspletosaurs have lachrimal horns with upper edges that are swollen side-to-side and bear coarse grooves and ridges. All the best preserved gorgosaur skulls — e.g. USNM 12814, AMNH 5336, 5664, FMNH 308 — have horns with apices that are displaced forward. Daspletosaurs and the new, unnamed tyrannosaur from the Horseshoe Canyon have horn apices that rise directly upwards. Tyrannosaurus and Tarbosaurus have no horn apices whatever and, instead, have dorsal-lateral bars of the lachrimal that are greatly swollen. In Tarbosaurus and Tyrannosaurus the postorbital part of the orbital rim is also greatly swollen, and, together the postorbital and lachrimal make a low, broad reinforced forehead area adapted for delivering head-butting blows. The apparently weak development of a rugose lachrimal bar or lachrimal horn in Nanotyrannus is a condition that cannot be derived easily from the lachrimal device seen in any other tyrannosaur. As we argue below, the ancestor of tyrannosaurs probably was an animal close in grade to a deinonychid and probably possessed a lachrimal with no horny development other than a slight thickening along the dorsal-lateral edge. Therefore the lachrimal condition in Nanotyrannus may well be the most primitive known for the Family Tyrannosauridae .
The ventral-anterior angle of the dorsal bar of the lachrimal in Nanotyrannus bears a depressed, smooth surface that forms the posteriondorsal corner of the antorbital fenestra ( Fig. 3 View Figure 3 ). The outer anterior edge of the vertical limb of the lachrimal bears a similar depressed surface that is continuous with the smooth, depressed zone that borders the ventral edge of the antorbital fenestra on the maxilla. All these depressed borders probably served as attachment areas for the anterior pterygoideus muscle bundles that bulged outwards through the fenestra. Nearly all theropods have a similar pattern. In Tyrannosaurus the entire preorbital part of the lachrimal is greatly swollen and the smooth depressed area has disappeared. In most theropods of a ceratosaur grade or higher, there is a pneumatic canal opening near the top of the posteriondorsal depressed area on the lachrimal. Nanotyrannus shows this condition. In Tyrannosaurus rex this foramen has become enclosed by swollen lachrimal bone on all sides, a condition also developed in Tarbosaurus but to a lesser extreme.
The nasals in Nanotyrannus are very narrow, as in all other Late Cretaceous theropods and in contrast with Jurassic taxa. All tyrannosaurs have snouts with some sort of rugosities on the dorsal surface of the nasals, but Nanotyrannus displays the weakest development of this character. Nanotyrannus shows only a set of shallow striae and ridges running fore-to-aft along the dorsal surface of the nasals ( Fig. 3 View Figure 3 , Plate 1-3 View Plate 1 View Plate 2 View Plate 3 ). Gorgosaurs, daspletosaurs, tarbosaurs and Tyrannosaurus show a much heavier ridge-and-furrow ornamentation that in life must have supported a very rough horny skin topography. The long-snouted Mongolian tyrannosaur Alioramus displays a grotesque set of oval hornlets on the nasal ( Fig. 11 View Figure 11 ).
Nanotyrannus agrees with other tyrannosaurs in having a large external foramen, opening upwards, in the jugal.
The antorbital fenestrae of Nanotyrannus are very large but very close in pattern to those of the gracile gorgosaurs ( Fig. 11 View Figure 11 ). As in all tyrannosaurids, there is a small anterior, accessory fenestra, a character that links tyrannosaurs with dromaeosaurs, troödonts and allosaurs. The snout, in lateral view, is longer and lower in Nanotyrannus than in any other tyrannosaurid except Alioramus and approaches the condition seen in dromaeosaurs.
Lower Jaw. In the type of Nanotyrannus the upper edge of the mandible and the lower teeth are hidden by the skull and upper teeth. On the outer surface of the mandible there is a clear line of separation between the dentarv in front and the surangular + angular behind ( Plate 1 View Plate 1 ). This line of separation shows no evidence of suturing or firm connection whatever, and undoubtedly there was a loosely articulating intramandibular joint as is found in all other tyrannosaurids. The intramandibular joint in tyrannosaurids is constructed very much like that of troödonts, dromaeosaurs, allosaurs and ceratosaurs ( Fig. 13 View Figure 13 ): the dentary and splenial enclose a spacious intramandibular cavity, open to the rear; the angular sends a forwardly directed prong into the cavity; the angular prong.
has no sutural connection to the inner walls of the intramandibular cavity and can twist and flex in all directions; the anterior edges of the surangular + prearticular have no firm suturing to the posterior edges of the splenial + dentary, so the intramandibular joint can permit a wide range of movement that could expand the gullet or adjust the pitch and position of the mandibular teeth relative to the upper teeth. The podokesaurid theropods of the Triassic and Early Jurassic lack an intramandibular joint and have, instead, a firm suturing of the posterior and anterior mandibular units. The Middle Jurassic megalosaurids probably lacked the joint also, but no complete megalosaurid lower jaw has been recovered. (We restrict the term “megalosaurid” to Megalosaurus bucklandi , Poikilopleuron bucklandi and closely allied species, as argued by Waldman, 1974).
Teeth. Nanotyrannus differs from all other tyrannosaurids, except possibly Alioramus , in retaining the primitive theropod condition of maxillary teeth that are compressed strongly from side-to-side, very wide front-to back relative to the crown height, and very sharp ( Fig. 12 View Figure 12 ). None of the teeth in the type of Nanotyrannus can be seen from the inner surface, but the three loose teeth in the Denver Museum collection seem to match quite closely with those in the middle and anterior sector of the maxilla of the type ( Fig. 12 View Figure 12 ). These loose teeth supply most of the information on tooth shape for the genus. In the loose tooth that is from the mid-maxillary row, the keel along the anterior edge reaches almost as far towards the crown base as does the posterior keel. The entire posterior keel is serrated but the basal part of the anterior keel is free of serrations. The anterior keel moves toward the inside of the tooth as the keel passes to the base; the posterior keel moves towards the outside. In the loose tooth that is from the front of the maxillary row, the crown is concave on the inner surface and the anterior keel is displaced further towards the inner edge. The only other Late Cretaceous theropod with maxillary teeth of a form similar to that of Nanotyrannus is the “Jordan theropod” described cy Molnar (19/8), an animal that may be a large dromaeosaur.
Gorgosaurs, daspletosaurs, Albertosaurus , larbosaurus and Tyrannosaurus all have maxi liar7 teeth with cross sections that are much thicker side-to-sde relative to their length fore-to-aft in the upper part of the crown, and the crowns are much taller relative to the anterior-posterior length ( Fig. 12 View Figure 12 ). The posterior maxillary teeth of Tyrannosaurus rex are very blunt and have an overall shape 1 i ke that of posterior teeth in Gorgosaurus and Daspletosaurus ; the crown crosssection is thick and nearly symmetrical. Clearly the function of these small teeth was different from that of the tall, strong anterior maxillary teeth — perhaps the posterior teeth were used to crack bones. In most of the other theropods — allosaurs, ceratosaurs, megalosaurs — the posterior maxillary teeth are shorter and blunter in the crown than are the anterior maxillary teeth, although the degree of heterodonty is not so marked as in T rex . In Nanotyrannus the posterior maxillary teeth are much sharper and more compressesd sideto-side than those in the same loci in the other tyrannosaurids.
The sharp, delicate, blade-like teeth of Nanotyrannus indicate that the bite delivered a long, narrow wound; surely this genus could not produce the bone-crushing bite of a tarbosaur or Tyrannosaurus . The tooth count in Nanotyrannus is 15 maxillary and 4 premaxillary, as in Gorgosaurus stembergi ; all other gorgosaurs have 13 maxillary teeth except for FMNH 308 which has 15 maxillary teeth. The high count in the latter specimen strongly suggests that it represents a species distinct from that of USNM 12814 and AMNH 5336.
The four premaxillary teeth in Nanotyrannus have outer (buccal) crown faces that are very narrow and crowded close together. The crown cross section must be very deep buccallingually and probably had a flat inner (lingual) face as in other tyrannosaurids. The inner edges of the teeth cannot be seen and the presence or absence of serrations cannot be determined. The first maxillary tooth has a form like that of the premaxillary teeth. All other gorgosaur species have a similar arrangement of incisiform teeth. Tarbosaurs and Tyrannosaurus have a large, massive first maxillary tooth. Incisiform premaxillary teeth with assymmetrical crowns are also present in ceratosaurids, allosaurids, dromaeosaurids (especially in Dromaeosaurus itself) and troödontids but are not developed in the primitive podokesaurs or megalosaurs. In all the theropods equipped with incisiform teeth, the right and left upper toothrows converge together to make a U-shaped symphysis as seen in dorsal or ventral view. Thus the incisor row is arrayed along a broad anterior arcade useful for cropping meat from bones or grooming skin and pelage. The podokesaurs and megalosaurs have V-shaped symphyses where the right and left tooth rows are nearly linear ( Waldman, 1974). Incisiform teeth in a U-shaped arcade and an intramandibular joint define the Neotheropoda ( Bakker, 1986). It is only among the tyrannosaurids that the premaxillary teeth become exceptionally crowded and very narrow across the buccal face.
The maxillary tooth row is nearly straight in Nanotyrannus and the teeth are set in the maxilla at right angles to the row, probably the primitive condition for the family. Some gorgosaurs and all daspletosaurs, albertosaurs, tarbosaurs and Tyrannosaurus have tooth rows that are more strongly convex-down, as seen in lateral view, and the anterior maxillary teeth tend to be procumbent ( Fig. 11 View Figure 11 ), Procumbent teeth suggest that the bite was delivered as the head and neck lunged forward at the parts of the prey’s body that were gently curved — such as the flank or belly. Procumbent teeth used in this way could bite a chunk of bone and flesh off the rib cage or abdominal wall. The sharp but delicate Nanotyrannus teeth, set vertically in the jaw, would be better designed for biting objects with less gently curved surfaces.
ANCESTRY AND EVOLUTION OF NANOTYRANNUS
In order to determine the primitive state of characters within the tyrannosaurids, we must know what groups are the sister clades of the family ( Fig. 2 View Figure 2 ). Dromaeosaurs differ from Jurassic dinosaurs and agree with tyrannosaurs, ornithomimids, troödontids and acrocanthosaurs and possible oviraptorids in possessing the derived characters of a tall, flat, wide ascending process of the astragulus and a dorsal column that is shortened relative to the pelvis. But dromaeosaurs are more primitive than the other groups in lacking extensive pneumatization of the otic region, and in retaining a primitive paroccipital process that is shallow top-to-bottom at its root. Therefore we take dromaeosaurs as the primitive sister group of the clade tyrannosaurs + troödonts + ornithomimids + oviraptorids + acrocanthosaurs. The four Late Cretaceous families — tyrannosaurs, oviraptorids, troödonts and ornithomimids — all share the derived character of a metatarsus that is more elongated and more compact from side-to-side than in acrocanthosaurs and thus probably comprise a clade with an origin in the early Late Cretaceous. Currie (1986) has shown that ornithomimids and troödonts share unusual basicranial characters and probably are more closely related to each other than either is to tyrannosaurs. From the foregoing analysis, it follows that for tyrannosaurs the most primitive sister group among Cretaceous theropods is the dromaeosaurs. Although troödonts and ornithomimids are closer cladistically to tyrannosaurids than are dromaeosaurs, in jaws and dentition and snout the dromaeosaurs probably are better guides for reconstructing the primitive tyrannosaurid condition. Omithomimids — ostrich dinosaurs — have muzzle and jaws very highly modified as toothless beaks and hence offer little clue to the primitive condition in tyrannosaurs. Troödonts are highly modified in a different direction — the teeth have relatively huge serrations on the trailing edge, unlike the more normal theropod serration pattern seen in tyrannosaurs. Dromaeosaurs show a basic, primitive theropod pattern of compressed, recurved teeth with fine serrations.
The teeth of Nanotyrannus are very similar to those of dromaeosaurs in degree of lateral compression. All other tyrannosaurs, with the possible exception of Alioramus , have maxillary teeth that are thicker side-to-side, and therefore, on dental evidence alone, Nanotyrannus could be argued to be the most primitive North American tyrannosaur..
The morphology of the Nanotyrannus skull displays several exceptionally advanced features, all or most of which are related to the development of stereoscopy and a sharper head-neck flexure. Indeed, these two functional shifts may be reflections of a single behavioral reorganization — by holding the muzzle flexed downwards, the reorientation of the eyes achieves the maximum overlap of visual fields. Great side-toside expansion of the basicranium and temporal regions also increases the size of the jaw-closing muscles and the leverage of the muscles flexing the neck laterally. In Tyrannosaurus the expansion of the temporal region occurs without constriction of the muzzle but in Nanotyrannus the muzzle has a very narrow width relative to the skull length. Theropod muzzles housed the anterior part of the pterygoideus D muscle, a very important jaw-closing unit, and the temporal expansion in Nanotyrannus could be, in part, a compensation for the decrease in muscle volume incurred in the snout.
The type of Nanotyrannus is the smallest adult tyrannosaurid skull known, only 572 mm long, fully 13% shorter than the types of Alioramus and Gorgosaurus stembergi (Table 1). And the degree of coossification in the dorsal temporal region in Nanotyrannus confirms that the type was fully adult, while, on the other hand, the types of G. stembergi and Alioramus remotus may well be subadult in size. Could Nanotyrannus evolve by a process of dwarfing from a more typical tyrannosaur of large size? Since Nanotyrannus is from the very last faunal age of the Cretaceous, and since all earlier tyrannosaurid skulls that are definitely adult are 40% larger, it is justifiable on stratophenetic grounds to deduce that the small adult size is a late-evolving derived character. On the other hand, as we argue elsewhere (Bakker, Currie, and Williams, in press), the most closely related sister group of tyrannosaurids are the oviraptorid, troödont and ornithomimid theropods, animals of small size, so a cladistic argument could be generated to defend the notion that small size is a primitive character for the Family Tyrannosauridae .
The apparently weak lachrimal horn of Nanotyrannus cannot be derived in any straightforward way from the low, grossly thickened lachrimal of Tyrannosaurus and Albertosaurus , or the more erect, pointed horn of Gorgosaurus and Daspletosaurus . Neoteny is a possible mechanism for producing a delicate adult horn from an ancestral state that was more swollen and more coarsely sculpted. However, the type of Gorgosaurus stembergi , although probably far from adult size, already has a swollen and rough-textured lachrimal horn. Lachrimal horns of various shapes have evolved many times independently among large theropods (but, strangely enough, never among theropods with skulls less than 300 mm long) and the primitive state for the Family Tyrannosauridae is probably a lachrimal like that of dromaeosaurs — without any erect horn and only slight thickening along the dorsallateral border (Bakker, Currie, and Williams, in press). At present we favor the hypothesis that the lachrimal condition of Nanotyrannus evolved from a hornless and unswollen ancestral state.
Whatever its evolutionary origins, Nanotyrannus seems to have evolved its stereoscopy independently of Tyrannosaurus . The low, long snout and primitive teeth of Nanotyrannus seem to preclude an ancestry from an animal as advanced as Gorgosaurus . The two stereoscopic genera, Nanotyrannus and Tyrannosaurus , are the only well known large predators from the Lancian Age, and it is noteworthy that both have achieved the highest degree of potential stereoscopy known among large theropods. Small theropods of the Cretaceous also show strong tendencies for frontal reorientation of the orbits — the orbits had moderate forward orientation in the Early Cretaceous dromaeosaurs and very strong forward orientation in ornithomimids and troödonts of the Late Cretaceous. Clearly natural selection was favoring greater precision of depth perception. A very similar trend towards greater stereoscopy can be seen among mammalian predators of the Cenozoic. The earliest predators of the Paleocene and Eocene — didymictids, miacids, hyaenodontids, oxyaenids — all had relatively small orbits that faced mostly outwards. But these archaic groups were replaced eventually by more modern cats, nimravids, dogs, bear-dogs, and hyaenas, all of which had relatively larger orbits with a greater degree of forward orientation. Thus Nanotyrannus seems to have been shaped by an inexorable law of the coevolution between predator and prey, a law that operated to produce ever greater finesse in the choreography of attack behavior.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nanotyrannus
| Bakker, R. T., Williams, M. & Currie, P. J. 1988 |
Gorgosaurus lancensis
| Gorgosaurus lancensis Gilmore, 1946 |
| Albertosaurus |