
Compsoneuriella thienemanni Ulmer, 1939
|
publication ID |
https://doi.org/10.11646/zootaxa.3835.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:5BBF2D80-F682-49E7-9B32-64E75ACAA6CF |
|
DOI |
https://doi.org/10.5281/zenodo.6124749 |
|
persistent identifier |
https://treatment.plazi.org/id/03A087A0-433C-4A06-FF7A-EB9A95C75923 |
|
treatment provided by |
Plazi |
|
scientific name |
Compsoneuriella thienemanni Ulmer, 1939 |
| status |
|
Compsoneuriella thienemanni Ulmer, 1939
Compsoneuriella thienemanni Ulmer 1939 , Arch Hydrobiol. Suppl. 16: p. 563 (adults), p. 672 (nymphs pro parte)
nec Compsoneuria thienemanni Braasch & Soldán, 1986 View in CoL , Reichenbachia 24(3), p. 41 (adult, nymph)
nec Compsoneuria thienemanni Boonsoong & Braasch, 2013 View in CoL , Zookeys 272, p.75 (nymph)
Material examined. Specimens in ethanol.
1 female imago, Sumatra, Wai Lima , Lampongs , XI–XII. 1921. Karny leg. [ ZMH]
2 male imagos (fore- and hind legs mounted on slide), 1 male subimago, Sumatra, Lake Singkarak , III.1929, Lampe , Prof. Thienemann leg. [ ZMH]
1 male subimago (with corresponding nymphal exuvia on slide), Sumatra, shore of the Lake Singkarak , 23.II.1929, F3e, Prof. Thienemann leg [ ZMH]
4 nymphs (one mounted on slide), Indonesia, West Java, Fish ponds south of the Puntjak pass , 13.IX.1928, B13, Prof. Thienemann leg [ ZMH, MZL]
1 nymph , Indonesia, West-Java, Buitenzorg, lateral branch of the Tjiliwung River , IX. 1928, B3, Prof. Thienemann leg [ ZMH]
3 nymphs (two mounted on slides), Indonesia, Sumatra Barat , Harau Canyon , stream, 540m, 0° 04.428’S 100° 38.002’E, 27.IX.2009, SUM09/UN11, M. Balke & D. Amran leg. [ ZMH, MZL] GoogleMaps
1 nymph, Indonesia, Sumatra Barat , Sawahlunto , Sikalang , 200m, 0° 37.678’S 100° 46.314’E, 21.IX.2009, SUM01, M. Balke & D. Amran leg. [ LIPI] GoogleMaps
Pinned specimen. 1 female imago, E. Jacobson, Toentang Java , Oct. 1910 [ ZMH] (sub. nom C. spectabilis )
Lectotype designation. Ulmer did not designate a holotype for Compsoneuriella thienemanni , although he mentioned that the type was a male imago; therefore all the original material constitutes the type series and consequently all specimens are syntypes. In the ZMH collections catalogue, under this species name, appears the mention of a holotype male imago. This information is also available in Weidner (1964). This terminology cannot be accepted because the “ holotype ” has not been designated by Ulmer and cannot be ascertained by the presence of a single specimen (see also Recommendation 73F. Avoidance of assumption of holotype, ICZN, 1999). Weidner (1962; 1964) also used the term “paratypoides” which should correspond to our actual concept of “ paratypes ”. Finally Ulmer himself brought some confusion; when describing the nymph of C. thienemanni , he mentioned in the material examined the four nymphs belonging to the species C. spectabilis (see above), with the following comment in bracket “darunter das typische Stück”, which means “among which the typical one”. This cannot be taken into account for the designation of a type specimen, and this is not a nomenclatorial act according to the Code.
As obviously all the specimens examined by Ulmer under the name Compsoneuriella thienemanni encompass more than one species, a lectotype is designated among them, which is the best preserved male imago, with dissected genitalia, and which bears the following labels:
i) Zool Mus. Hamburg—Coll. G. Ulmer—Eing Nr. 6–1963
ii) Compsoneuriella Thienemanni Ulm.
iii) Singkarak, Lampe III. 29;
iii) Compsoneuriella thienemanni Ulmer, 1939 LECTOTYPUS ♂, M. Sartori des., 2013
The type locality is therefore restricted to Singkarak, Sumatra, Indonesia (articles 73.2.3 and 76.2, ICZN, 1999).
Description. Male imago ( lectotype). Coloration faded, legs yellowish, thorax and compound eyes brownishorange, wings translucent, costal and subcostal fields milky in distal part, abdomen translucent, except segments VII–X yellowish, with ventral nerve cord whitish, gonopods yellowish, cerci whitish, some junctions between segments lightly tinted in orange in proximal part, suggesting cerci originally banded with darker color on ca. onethird of their length.
Measurements: Body length: ca. 6.6 mm; Forewing length: 6.3 mm; Gonopods length: 0.8 mm; Cerci length: ca. 10 mm but probably broken at apex.
Head: Compound eyes large, meeting in middle at distance superior to distance between lateral ocelli; lateral ocelli two times larger than median ocellus. Frons translucent with well-marked carina.
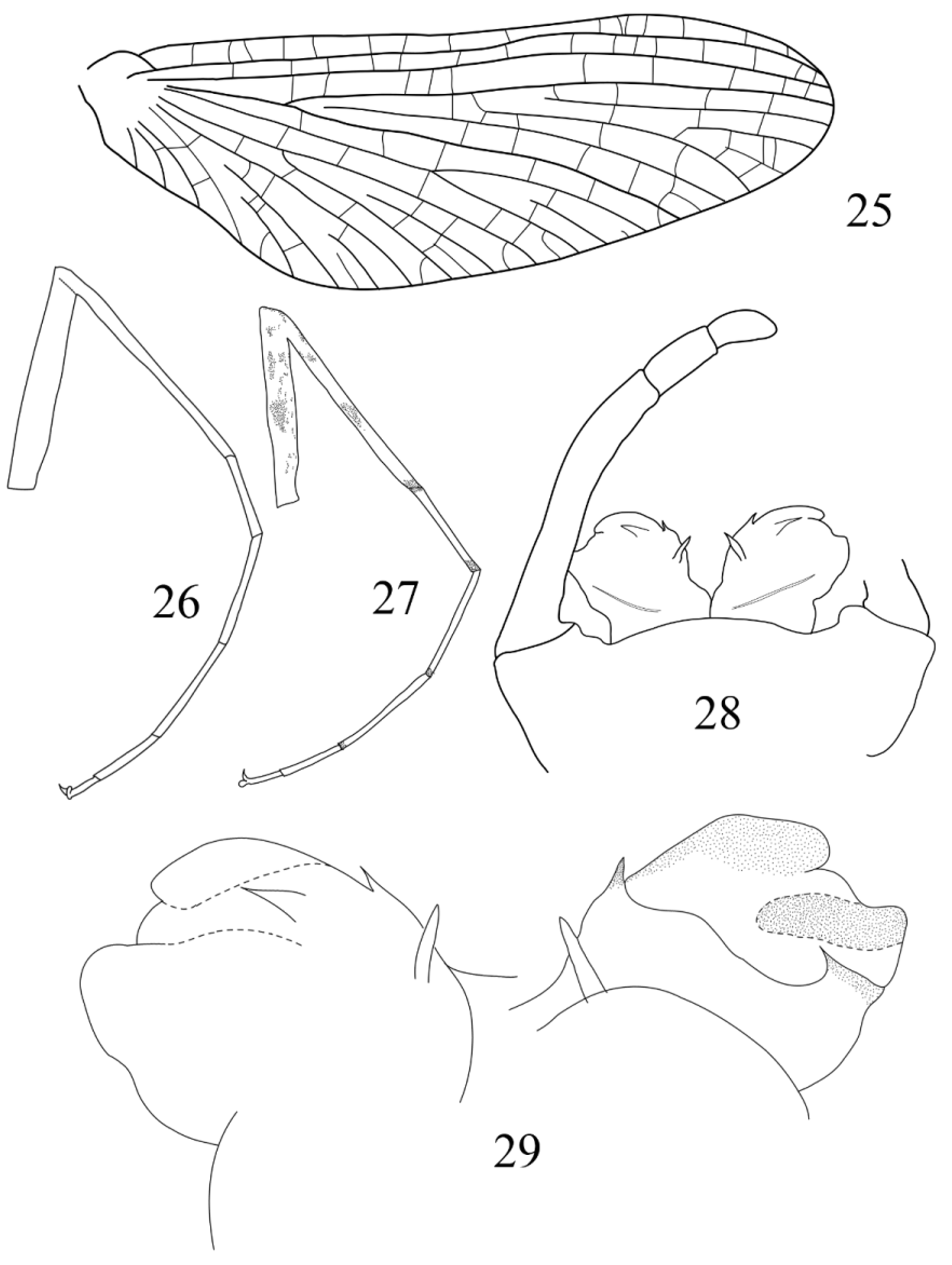
Thorax: Transversal suture on mesothorax present; medial depression of furcasternum of mesothorax subparallel. Forewing ( Fig. 25 View FIGURES 25 – 29 ) with numerous crossveins, those of costal and subcostal fields tinted with orange (blackish originally?); no rows of crossveins visible. Foreleg ( Fig. 26 View FIGURES 25 – 29 ) with tibia slightly longer than femur, and tarsi 1.75x longer than tibia; tarsal segment 2 and 3 subequal in length, segment 1 shorter, ca. 0.7x length of segment 2, segment 4 0.55x length of segment 3 and segment 5 0.5x length of segment 4; tarsal claws dissimilar, one obtuse, one hooked. Hind leg with tibia shorter than femur, tarsi subequal length of tibia, tarsal segments 1 longest, segments 2, 3 and 5 subequal, only slightly shorter than segment 1, segment 4 shortest.
Abdomen (genitalia): styliger plate ( Fig. 28 View FIGURES 25 – 29 ) slightly convex; basal segment rounded, 2x longer than wide, with inner corner well-marked; segment 1 short and broad, segment 4 slightly shorter than segment 3, together between ½ and ⅓ length of segment 2. Penis lobes stocky. In ventral view ( Fig. 29 View FIGURES 25 – 29 , left), apical sclerite reaching half of lobe width, with distinct basal spine; well-developed subapical spine present below apical sclerite; on inner face of lobes, two stout and pointed titillators. In dorsal view ( Fig. 29 View FIGURES 25 – 29 , right), lateral sclerite narrow and constricted at apex, basal sclerite strongly developed and bearing on outer margin one small and acute spine directed inwards.
Nymph. Size: body length up to 7mm and 8 mm in male and female nymphs respectively.
Coloration ( Fig. 81 View FIGUR ): overall medium brown with yellowish brown markings. Head ( Fig. 30 View FIGURES 30 – 35 ) with lateral margin yellowish and 4 rounded spots near anterior margin, median spots apparently extend backward in yellowish band through antennae and to anterior margin of compound eyes. Femora also contrasted, yellowish with two brownish maculae longitudinal to outer margin and two to inner margin, base of stout bristles on upper face strongly colored in dark brown; tibiae whitish with three ringed brownish maculae, largest macula in middle of tibia, others present in proximal and distal positions. Prothorax greyish brown with two diagonal elongated yellowish bands on each side, mesothorax with two blackish maculae situated medially and close to prothorax. Abdominal tergites with much contrasted patterns: tergite III with two median and longitudinal black thick lines, tergite IV with triangular black lines not reaching posterior margin of segment, tergite V with small triangular black lines on proximal fourth of segment, tergite VI with blackish lines faded in background, but with two rounded yellowish anterior dots, tergite VII broadly similar to tergite IV, tergites VIII and IX dark brown with two small yellowish dots in middle of segment, tergite X medium brown with two large yellowish maculae in middle of segment, reaching anterior margin. Cerci light brown, banded with dark brown segments.
All patterns slightly variable from one individual to another, with respect to size and intensity of maculae, black lines and spots.
Head: Labrum ( Fig. 31 View FIGURES 30 – 35 ) greatly extended laterally, with narrow tips; anterior margin almost straight, covered by numerous long and simple setae; dorsal surface with long row of small stout setae; ventral surface with scattered long and simple setae directed toward middle of labrum and concentrated near anterior margin. Mandibles strongly arched, both with lateral margin covered with several rows of long and thin setae; right mandible ( Fig. 32 View FIGURES 30 – 35 ) with outer incisor saw-like on both margins (those on outer margin often difficult to see on microscopic slides, Fig. 32 View FIGURES 30 – 35 c), composed of ca. 6 teeth, inner incisor trifid ( Fig. 32 View FIGURES 30 – 35 b); presence of 4 fimbriate setae below incisors and 4 long and simple setae below mola ( Fig. 32 View FIGURES 30 – 35 a); left mandible with outer incisor saw-like on its inner margin, composed of ca. 6 teeth, and with subapical tooth in ventral position; inner incisor slender and pointed, bearing 2 setae-like structures in apical position ( Fig. 33 View FIGURES 30 – 35 b); 3 fimbriate setae present below incisors, followed by tuft of short setae above mola, and 5 long and simple setae below mola ( Fig. 33 View FIGURES 30 – 35 a). Maxillae with row of ca. 12 comb-shaped setae on anterior margin, medial ones bearing 7–9 teeth ( Fig. 54 View FIGURES 48 – 54 ); ventral surface of galea covered by scattered long fimbriate setae ( Figs 52–53 View FIGURES 48 – 54 ); proximal dentiseta bifid and fimbriate, with two distal dentisetae simple ( Figs 48–51 View FIGURES 48 – 54 ). Hypopharynx ( Fig. 34 View FIGURES 30 – 35 ) with well-developed superlinguae, densely covered with long and simple setae on lateral margin, ending below rounded apex of lobes; lingua without or with few short setae arranged in tuft. Labium ( Fig. 35 View FIGURES 30 – 35 ) with glossae somewhat slender and with inner margin slightly concave before apex, inner margin covered with dense simple setae, outer margin with fimbriate setae, dorsal face with single pointed bristle; paraglossae large and extending laterally, two times longer than wide.
Thorax: Leg ornamentation rather similar between all three pairs, except forefemora lack row of stout spines on inner margin. Supracoxal spurs strongly developed, especially on mid- and hindlegs, acute apically. Hindleg ( Figs 36 View FIGURES 36 – 40 , 64 View FIGURES 60 – 67 ) with femur bearing row of long and simple setae on its outer margin, together with stout spines on distal half; inner margin with row of stout spines directed toward apex, row of long and very thin setae and incomplete submarginal row of stout spines ( Fig. 36 View FIGURES 36 – 40 ); bristles on upper face of femora elongated, with lateral margins clearly convergent, with rounded apex ( Figs 37 View FIGURES 36 – 40 , 66 View FIGURES 60 – 67 ); tibiae ( Fig. 65 View FIGURES 60 – 67 ) each with one row of long and very thin setae on outer margin; inner margin with only 5–6 short spines regularly arranged. Tarsal claw ( Fig. 67 View FIGURES 60 – 67 ) hooked, with no subapical denticles.
Abdomen: Posterolateral projections weakly developed on segments I–III, more pronounced on segments IV–IX. Gills ( Fig. 38 View FIGURES 36 – 40 ) broadly oval or rounded and asymmetrical, fibrillate part well developed on gills I–VI, absent on gill VII; plate gills with tracheation apparent, gill VII ( Fig. 39 View FIGURES 36 – 40 ) more elongated than others, with inner margin concave near apex. Posterior margin of tergites ( Fig. 40 View FIGURES 36 – 40 ) with row of large and pointed spines, together with intercalary small and medium ones; several rows of microdenticles apparent. Cerci with whorl of stout and pointed setae on each segment.
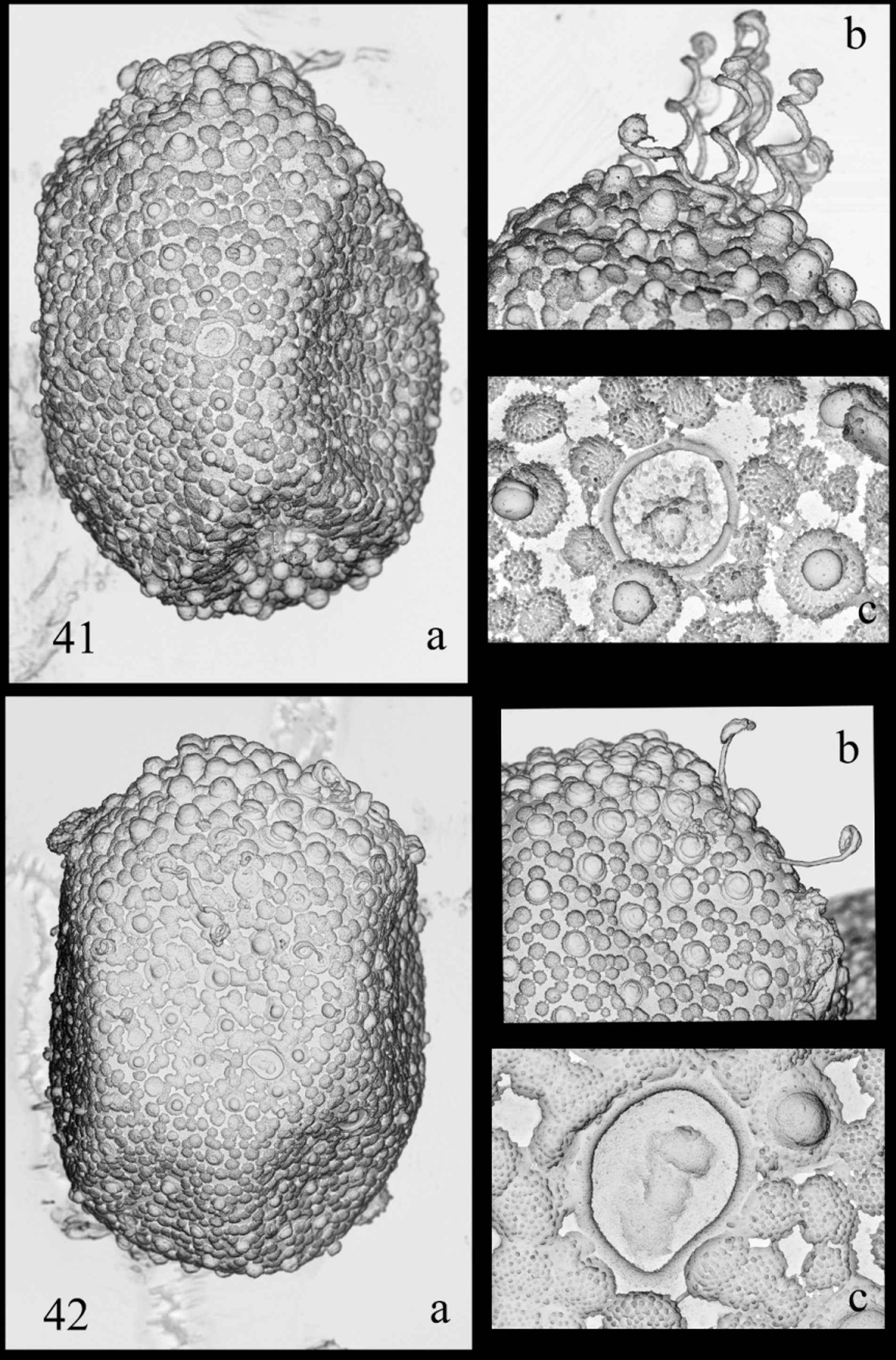
Eggs (from mature nymph and female): Size: 110–125 µm x 85 –100 µm. Chorion covered with flat, fibrillose and rounded structures ( Figs 41 View FIGURES 41 – 42 a, 42a), some bearing KCTs; on each pole, KCTs around two times size of KCTs located elsewhere ( Figs 41 View FIGURES 41 – 42 b, 42b); micropyle with single margin ( Figs 41 View FIGURES 41 – 42 c, 42c).
Discussion. The lectotype description fits the diagnosis proposed by Ulmer (1939), but there are some intriguing points. The first one concerns the forewing venation. When describing its new genus Compsoneuriella, Ulmer stated that the wing venation was very close to the one of Compsoneuria (…”ganz ähnlich wie bei Compsoneuria Etn ”), a statement not correct when comparing our Figs 1 View FIGURES 1 – 7 and 8 View FIGURES 8 – 12 , with Ulmer´s (1939) fig. 143 and Ulmer´s (1924) fig. 55. In C. thienemanni , the arrangement and number of crossveins are quite different than in C. spectabilis . The “rows” of crossveins are absent, and the crossveins are more numerous than in the previous species, especially in the costal and subcostal fields. An answer to this “paradox” can be found in his 1924 paper. When redescribing C. spectabilis , he illustrated the forewing ( Ulmer 1924, fig. 54) and the abdomen in lateral view ( Ulmer 1924, fig. 53B) of a pinned female imago (coll. Jacobson), mentioning that this female possesses different abdominal color patterns as well as different wing venation, but Ulmer considered them as intraspecific variation. When making the diagnosis of the genus Compsoneuriella ( Ulmer 1939, p. 563), he wrote that the wing venation of the new genus is similar to the one of this female from 1924 (“wie bei der von mir in Treubia, 6, 1924, f. 53b, 54 abgebildeten dunkleren Form (Tuntang, Java) der C. spectabilis Etn ”). This female possesses numerous crossveins, more than 15 in the costal field and more than 12 in the subcostal one, and the general pattern is similar to the lectotype of Compsoneuriella thienemanni . One can therefore conclude that this female was misidentified by Ulmer, and rather than being a variation of C. spectabilis , constitute a female imago of C. thienemanni .
The foreleg proportions illustrated in my Fig. 26 View FIGURES 25 – 29 are a little bit different from the drawing made by Ulmer (1939, fig. 140) as well as his description, and this can be due to some distortion in the drawing since it has been done with the leg attached to the body, and therefore all pieces not always in the same plane. Fig. 27 View FIGURES 25 – 29 presents the leg of a specimen on Ulmer’s slide, which can belong to the lectotype or to the other male imago, and which kept its original coloration. In this figure, the leg proportions are more in accordance with Ulmer’s description, except that tarsal segment 3 is longer than the first one, whereas Ulmer stated that they are of the same length. The coloration of the leg is as follow: on the dorsal face of the femur, the same small blackish maculae as in C. spectabilis can be found; on the ventral side there are three large dark spots, one apical, one subapical and one in the middle of the femur; tibia is ringed by three maculae, one apical, one in the middle and one subproximal; distal part of the tarsal segments tinted in blackish, the coloration fading towards the apex of the tarsus to become light brown on segment 4.
Major differences have to be found on the genitalia. Ulmer’s drawings (1939, figs. 145–149) did not mention any spines or sclerotized structures, but the general shape is in accordance with the drawings. These genital structures are difficult to represent because the penis is thick due to a very developed basal sclerite on dorsal side, and the structures not easy to see. Anyway, penis lobes bear a spine of the apical sclerite together with subapical spine below the apical sclerite, an inner spine on the basal sclerite, and titillators. Although broadly similar, genitalia of C. thienemanni differ from those of C. spectabilis in several respects; the basal sclerite is much more developed and the basal spine is small, the lateral sclerite is narrower, the apical spine is less developed and the titillators are much more developed than in C. spectabilis .
One misinterpretation by Ulmer needs to be corrected. When describing the maxillae, he mentioned that the crown of the galea bears two rows of comb-shape setae ( Ulmer 1939, fig. 453e); in fact the mounted nymph was about to molt, and what Ulmer interpreted as a second row is in fact the row of the next molt. The nymph of C. thienemanni described here is very closely related to the one illustrated by Braasch & Soldán (1986a) under the name Compsoneuria thienemanni (?) from Vietnam, and reported later on also from Thailand ( Braasch & Boonsoong 2010). In peculiar the abdominal patterns are almost identical between insular and continental populations (see Braasch & Soldán 1986a, fig. 14.5 and Boonsoong & Braasch 2013, fig. 3I). Anyway, both are distinct species, as demonstrated below.
Compsoneuriella thienemanni occurs in Java and Sumatra; there is no evidence at the moment that the species inhabits other islands. The pinned specimens reported by Ulmer (1939, p.566) from Borneo (Nanga Serawei, 12- 18.XI.1924, Prof. Winkler leg) could not be related to C. thienemanni , although probably belonging to the genus Compsoneuriella (see below).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Compsoneuriella thienemanni Ulmer, 1939
| Sartori, Michel 2014 |
Compsoneuria thienemanni
| Boonsoong & Braasch 2013 |
Compsoneuria thienemanni Braasch & Soldán, 1986
| Braasch & Soldan 1986 |
Compsoneuriella thienemanni
| Ulmer 1939 |