
Cyrtodactylus vairengtensis, Lalremsanga & Colney & Vabeiryureilai & Malsawmdawngliana & Bohra & Biakzuala & Muansanga & Das & Purkayastha, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5369.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:030933EB-07C2-48BE-9B7F-EEACFA102A50 |
|
DOI |
https://doi.org/10.5281/zenodo.10248411 |
|
persistent identifier |
https://treatment.plazi.org/id/039B87C5-466C-FF8D-93B1-FF2AFF414540 |
|
treatment provided by |
Plazi |
|
scientific name |
Cyrtodactylus vairengtensis |
| status |
sp. nov. |
Cyrtodactylus vairengtensis sp. nov.
Figs. 3–7 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 ; Table 3 View TABLE 3
Holotype: Adult male ([ MZMU 2903 ]; Fig. 3 View FIGURE 3 ), from the jointed weathered siltstone bed along a public link road (24.501111°N; 92.762222° E; elevation 230 m a.s.l.), in the backyard of UPC-NEI church building, Hall Veng , Vairengte , Kolasib District, Mizoram, north-east India collected on 5 June 2022 by Zosiamliana Colney and Joseph Vanlalhriata. GoogleMaps
Paratypes: One adult male ( MZMU2904 ) and one female ( MZMU2905 ), same collection details as holotype . Two adult males ( MZMU2901 , MZMU2902 ) collected from a rocky wall covered by vegetation along Aitlang road (24.495556 °N; 92.757500 °E; elevation 168 m a.s.l.), Field Veng , Vairengte, Kolasib District, Mizoram, north-east India on 5 June 2022 GoogleMaps by Zosiamliana Colney, H. T. Lalremsanga, Lal Muansanga and Joseph Vanlalhriata and two adult females ( MZMU2626 , MZMU2742 ) collected along a narrow public slope stone steps walkway (24.501111 °N; 92.762222°E; elevation 230 m a.s.l.), Hall Veng , Vairengte , Kolasib District, Mizoram, northeast India by Zosiamliana Colney, C. Lalfamkima, Alfred Lalsanglura and Joseph Vanlalhriata GoogleMaps on 12 November 2021 and 2 April 2022.
Diagnosis: Cyrtodactylus vairengtensis sp. nov. is a moderate-sized gecko (adult SVL 57.6–73.6 mm) having 10–11 supralabials and 10–11 infralabials; 22–23 longitudinal rows of dorsal tubercles which are rounded and conical to feebly keeled; 34–39 continuous series of paravertebral tubercles between the level of axilla and groin; 35–41 mid-ventral scale rows across the ventral region; 9–11 precloacal pores in males; 5–9 precloacal pits (no pitted scales in one specimen) in females and 14–17 subdigital lamellae under toe IV; ventral surface of tail without transversely arranged enlarged subcaudal scales. Dorsum has dark brown blotches intermixed with bordering whitish patches that form an irregular body pattern; original tail has alternating dark brown and transverse whitish bands.
Description of holotype: Holotype in good preservation condition. Adult male, SVL 60.6 mm. Head less than one-third of SVL (HL/SVL 0.30), longer than broad (HW/HL 0.55), somewhat depressed (HD/HW 0.68) and distinct from neck, snout less than half of the head length (SO/HL 0.36); scales on the canthus rostralis, snout and forehead are homogeneous, interorbital area flat, canthus rostralis broadly rounded; loreal region inflated with granular scales; scales on the interorbital and occipital regions somewhat homogeneous in size; scales on the interorbital region of the head to the nape are slightly smaller than those of the forehead, somewhat blunt and juxtaposed having no distinct tubercles; eye approximately one-quarter of the head length (OD/HL 0.23); vertical pupil with crenulate margins; supraciliaries moderate in size, blunt, somewhat oval, those present in the middle (above the orbit) are largest and most prominent; ear opening oval, obliquely orientated, small (EL/HL 0.077); orbit to ear distance is less than the eye diameter (OE/OD 0.93); rostral wider than long (RL/RW 0.57), partially and dorsally divided by a poorly developed rostral groove; single enlarged supranasal on either side, separated by five small granular internasals; rostral in contact with the first supralabial, nasals, two supranasals and five small internasals; nostrils semicircular, openings laterally orientated, posterior half covered by the nasal pad, each nasal is in broad contact with the rostral and surrounded by the supranasal, first supralabial, and four small postnasals; a single row of small granular scales separate the orbit from the supralabials; mental wider than long (ML/MW 0.53), triangular; two well developed postmentals on either side; the inner pair of postmentals more than one and a half of the size of the outer pair (PMIIL/PMIL 0.52), bordered by the mental, infralabial I, the outer postmental and five gular scales on either side; outer postmentals bordered by inner postmental, infralabials I and II, and four gular scales on either side; 11 supralabials on each side, bordered by a row of medium to large sized scales, somewhat flat and elongated; 11 infralabials on each side, a row of enlarged gular scales (largest anteriorly) ventrally bordering infralabials II to V; gular region with small granular scales throughout except for a few scale rows bordering the mental, postmentals and infralabials which are larger, flat and juxtaposed.
Body moderately slender, trunk length approximately half of the snout to vent length (TRL/SVL 0.47); dorsal scales are mostly heterogeneous, small rounded granule like scales intermixed with irregularly arranged, bluntly conical and feebly keeled enlarged tubercles (3–4 times the size of granular scales) becoming slightly smaller and more conical towards the flanks, the largest on the mid-dorsum; tubercles extending posteriorly from the occipital region to the second segment of the tail and are smaller on the nape than those of the dorsum; 23 mid-dorsal tubercle rows throughout the dorsum; 34 paravertebral tubercles between the level of the axilla and the level of the groin; ventrolateral folds are poorly developed, not denticulate but with a single row of continuous, projected smooth tubercles; ventral scales smooth, cycloid, imbricate to sub-imbricate and are in 40 rows, much larger than dorsals and are slightly smaller in size under thighs; ten distinct precloacal pores in a continuous series; a single row of six slightly enlarged scales (median scale being the largest) between the precloacal pores and the vent; four small postcloacal tubercles on both the sides of the tail base. Forearm (FL/SVL 0.16) and tibia (CL/SVL 0.18) short; digits narrow, without a scansorial pad, strongly inflected at each joint, all bearing robust, recurved claws; subdigital lamellae transversely widened beneath the basal phalanx; basal lamellae 4-5-6-5-5 on the right manus and 4-6-5-6-5 on the right pes; distal lamellae (intervening rows of non-lamellar granular scales between the basal and distal lamellae series in parentheses): 5(4)-7(2)-8(3)-8(3)-8(3) on the right manus, 5(4)-7(2)-10(2)-8(4)-10(3) on the right pes; interdigital webbing absent from both the manus and pes; relative length of digits: I <II <V<III<IV on the right manus, I <II <V<III<IV on the right pes; scales on the palms and soles are smooth, weakly raised, subimbricate; scales on the forelimbs are heterogeneous in size, comprising flat, imbricate to subimbricate scales on the upper arms, and those on forearms are heterogeneous in size, ventral portion covered with heterogenous sized imbricate scales; scales on the hindlimbs are heterogeneous in size, dorsal surfaces of the thighs and shanks have slightly enlarged scales, intermixed with scattered, enlarged, conical, feebly keeled tubercles; anterior portion of thighs and ventral aspect of hindlimbs have slightly enlarged, somewhat smooth, imbricate to subimbricate scales.
Tail complete, oval in cross-section, dorsoventrally depressed; dorsal tubercles on the tail base (tail portion containing the hemipenes) are enlarged, flat, and pointed; few enlarged, flat and distinctly keeled, tubercles randomly scattered near the first segment of the tail gradually decreasing slightly towards the second segment, remaining dorsal caudal scales smooth, flat, heterogenous in size and shape; subcaudal scales smooth, imbricate and heterogeneous in size, forming a mid-ventral series of granular scales; no enlarged transverse or paired row of subcaudals.
Colouration in life ( Fig. 4A View FIGURE 4 ): Dorsum primarily greyish brown in colour; head somewhat brownish grey; forehead without markings; nape with a pair of discontinuous longitudinal markings, somewhat parallel to each other. A distinct, broad, dark-edged post ocular stripe with a lighter border runs from behind eyes to ear opening. Dark brown blotches with lighter edges distributed across dorsum, sometimes forming a reticulate pattern. Forelimb and hindlimb with indistinct whitish or cream-coloured blotches intermixed with dark brown blotches. Tail with alternating dark brown and transverse white bands. Venter off white.
Colouration in preservative: The colour is pale in comparison to the live specimen and the dark spots on the dorsum have partially darkened.
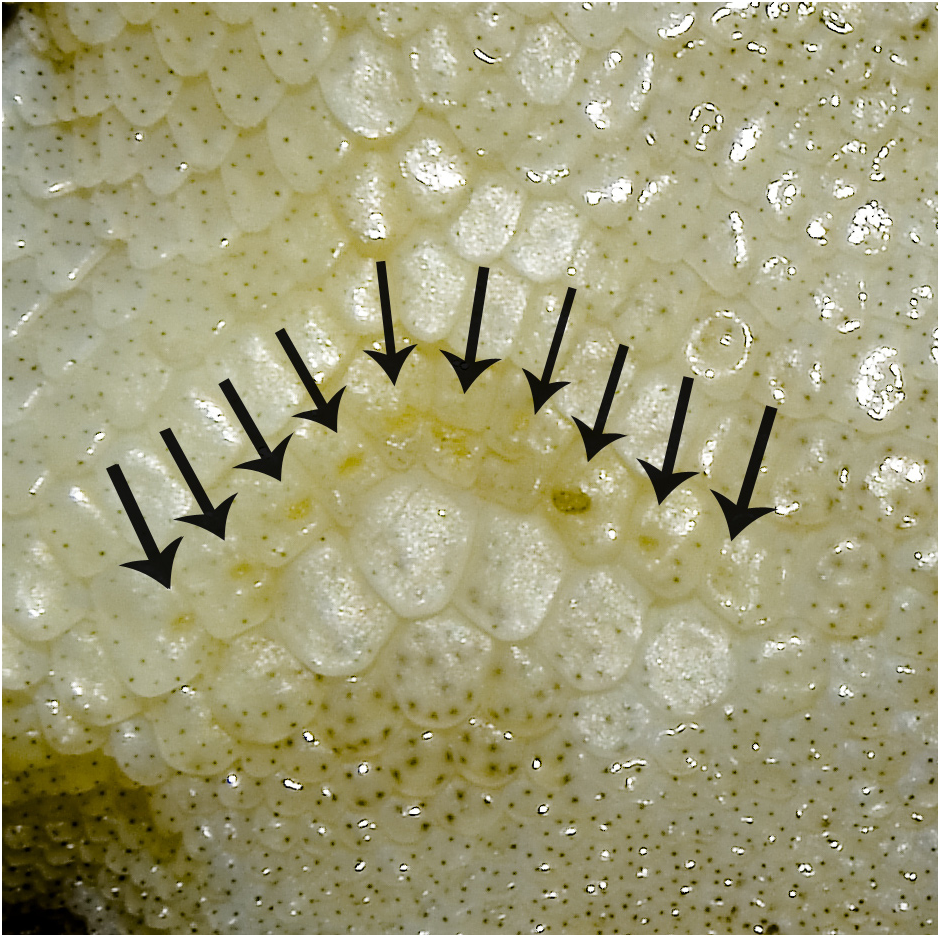
Variation: Refer to Table 3 View TABLE 3 for meristic, morphometric and basic pholidosis variation within the type series of C. vairengtensis sp. nov. ( Fig. 5 View FIGURE 5 ). The type series of C. vairengtensis sp. nov. includes three adult males (MZMU2901, MZMU2902 and MZMU2904) having prominent pre-cloacal pores (9–11) and three adult female specimens (MZMU2905, MZMU2626, and MZMU2742), all having visible pitted scales (5–9) in their precloacal region except MZMU2742 have none.
Comparisons: Here, we have compared Cyrtodactylus vairengtensis sp. nov. with all described members of the Cyrtodactylus khasiensis clade, as well as with C. kamengensis of the C. peguensis clade including species ( C. cayuensis , C. martinstollii , C. markuscombaii , C. himalayicus , C. tamaiensis ) that are not assigned to any group/ clade due of the absence of molecular data.
We have differentiated Cyrtodactylus vairengtensis sp. nov. from its congeners by a combination of the following characters: a smaller maximum adult size, SVL 73.6 mm max. ( C. kazirangaensis min. 80.0 mm; C. montanus min. 78.2 mm; C. arunachalensis max 81.7 mm; C. khasiensis max. 81.1 mm; C. martinstolli max. 82.0 mm; C. tamaiensis max. 90.0 mm; C. cayuensis max. 79.8 mm; C. jaintiaensis min. 96.2 mm; C. brevidactylus max. 88.0 mm; C. chrysopylos max. 83.8 mm; C. aunglini max. 81.6 mm); presence of 9–11 distinct precloacal pores in males ( Fig. 7 View FIGURE 7 ) versus 6 PcP in C.ngopensis , 12 PcP in C.namtiram , 3–5 PcP in C. lungleiensis , 5–7 PcP in C. bengkhuaiai , 7–8 PcP in C. aaronbaueri , 11–18 PcP in C. agarwali , 34–38 PcFP in C. karsticola , 40 PcFP in C. tamaiensis , 26–39 PcFP in C. guwahatiensis , 7 PcP in C. markuscombaii , 14 PcP in C. septentrionalis , 29–37 PcFP in C. tripuraensis , 12–13 PcP in C. aunglini , 0–8 PcP in C. martinstolii , 16–29 PcFP in C. gansi and 7 PcP in C. siahaensis ; 5–9 precloacal pits (no pitted scales in one specimen) in females versus no visible pitted scales in females of C. bengkhuai , 0–3 pitted scales in C. siahaensis and 10–13 (no pitted scales in 1 specimen) precloacal pits in C. bapme ; 22–23 rows of feebly keeled, conical dorsal tubercles throughout the dorsum versus 19–20 rows in C. ngopensis , 24–26 rows in C. arunachalensis , 21 rows in C. namtiram , 18 rows in C. cayuensis and C. mandalayensis , 14–15 rows in C.markuscombaii , 16–18 rows in C. nagalandensis , 16–20 rows in C. chrysopylos , 21 rows in C. tamaiensis , 24–28 rows in C.lungleiensis , 23–27 rows in C. mombergi , 27–30 rows in C. brevidactylus , 19–20 rows in C. jaintiaensis , 19-21 rows in C. tripuraensis and 21 rows in C. himalayicus ; presence of 34–39 rows of paravertebral tubercles between the level of axilla and groin separates it from C. exercitus (32–34 rows) C. jaintiaensis (30–34 rows) and C. septentrionalis (38–42 rows); 35–41 rows of mid ventral scales across the venter (32 rows in C. mandalayensis ; 30–34 rows in C. kamengensis and C. urbanus , 30–35 rows in C. guwahatiensis ; 57 rows in C. myaleiktaung ; 34- 35 rows in C. nagalandensis ; 33 or 34 rows in C. himalayicus and 47–49 rows in C. aunglini ); 14–17 subdigital lamellae beneath fourth toe (versus 9–13 subdigital lamellae in C. kamengensis , 18–19 in C. brevidactylus , 19–23 in C. aunglini and C. chrysopylos ; 13 in C. martinstolli ; 18 in C. myaleiktaung , 19 in C. markuscombaii ; 19–22 in C. khasiensis and C. mombergi ; 10 in C. himalayicus and C. gansi ).
A single row of 5–6 slightly enlarged scales below the pored scales (median scale being the largest) distinguishes Cyrtodactylus vairengtensis sp. nov. from C. arunachalensis , which has three to four rows of slightly enlarged scales below the pored scales. Moreover, the presence of indistinct dark dorsal spots in the dorsum differentiates Cyrtodactylus vairengtensis sp. nov. from C. bapme (7-9 paired dark blotches), C. urbanus (6–8 paired dark brown blotches in the dorsum), C. agarwali (8–9 pairs of dark blotches in the dorsum), C. aaronbaueri (11–12 prominent blotches in the dorsum), C. ayeyarwadyensis (9–11 paired dark blotches in the dorsum) and C. tripuraensis (presence of dark brown blotches in the dorsum usually bordered posteriorly by white punctuations in the latter two species), as well as from C. khasiensis , C. martinstolii and C.cayuensis by the presence of uniform granular subcaudals (versus presence of a median series of paired enlarged subcaudals in the latter three species).
In Cyrtodactyus vairengtensis sp. nov., the dorsal tubercles reach at least the second segment of the tail, which differentiates it from C. kazirangaensis as well as from C. montanus (tubercles not extending beyond first tail segment and tubercles reaching at least till the third tail segment respectively in the latter two species).
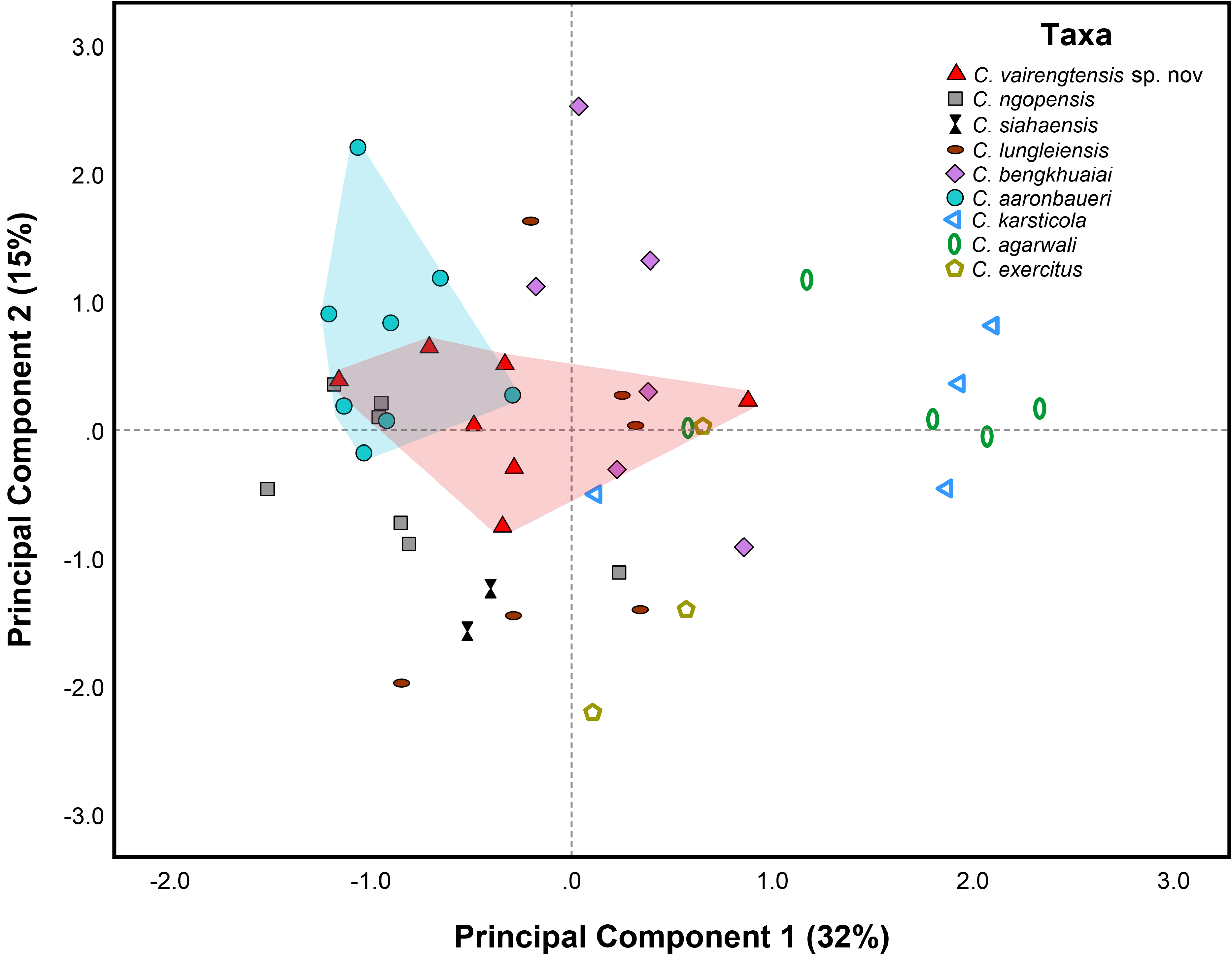
We performed statistical analyses based on the meristic values ( Table 3 View TABLE 3 ) to compare the new specimens with respect to the recently described ‘Mountain clade’ species from Mizoram state ( C. ngopensis , C. aaronbaueri , C. bengkhuaiai , C. lungleiensis , C. siahaensis ) along with certain members of the ‘Lowland clade’ from Meghalaya state ( C. agarwali , C. karsticola , C. exercitus ). In males, one-way ANOVA with taxon as a factor showed significant differences in the PcP/PcFP (F 8, 22 = 113.56, p <0.001), IL (right) (F 8,22 = 3.41, p = 0.011); FILamc (F 8,22 = 3.52, p = 0.009), FIILamb (F 8,22 = 2.44, p = 0.047), FIIILamc (F 8,22 = 5.12, p = 0.001), TILamc (F 8,22 = 3.16, p = 0.016), TIIILamb (F 8,22 = 2.76, p = 0.028), and TIVLamb (F 8,22 = 2.42, p = 0.048); and through the alternative Kruskal-Wallis H test, significant differences are also seen in the FIVLamc [H (8) = 21.59, p = 0.006], TILama [H (8) = 17.09, p = 0.029], TIILamc [H (8) = 20.47, p = 0.009], and TIVLamc [H (8) = 15.67, p = 0.047]. In females, one-way ANOVA with taxon as a factor also showed significant differences in the pitted precloacal scales (F 5,11 = 3.24, p = 0.049), PVT (F 5,11 = 3.60, p = 0.036), DTR (F 5,11 = 25.36, p <0.001), and TILamc (F 5,11 = 8.70, p = 0.002). Excluding the sex-specific meristic characters (i.e. PcP/ PcFP in males and pitted precloacal scales in females), the interspecific different characters identified through univariate analyses among males (IL (right), FILamc, FIILamb, FIIILamc, FIVLamc, TILama, TILamc, TIILamc, TIIILamb, TIVLamb, and TIVLamc) and females (PVT, DTR, TILamc) were further adopted for determining the clustering of the species through multivariate analysis (PCA) by pooling sexes ( Fig. 8 View FIGURE 8 ). Given that the correlation matrix showed weak correlations between the variables (r <0.75), we utilized all the selected variables for the PCA. A total of five components were extracted (eigen values> 1) that accounted for 75% of the total variation, and the three components accounted for 59% of the total variation of the data, with PC1, PC2 and PC3 representing 32%, 15%, and 11% respectively. The representation of the first two components depicts the clustering of the new specimens and C. aaronbaueri with a marginal separation between them along the first and second principal components. Furthermore, ordination of the second and third components also depicts a marginal separation between the new specimens and C. aaronbaueri along the second principal components. Therefore, based on morphology and genetic assessment, we consider the new specimens from Vairengte as a new species of Cyrtodactylus .
Distribution and Natural History: We have collected individuals of the new species only from Vairengte town, one of the townships in the Kolasib District, bordering the Cachar District (Assam) in the extreme northnortheast of Mizoram. The area lies with in an ecotonal region between the Lushai upland hills in Mizoram and the Cachar lowland plains in Assam. The approximate aerial distances from the type locality of the new species to the type locality of its closest sister species, C. aaronbaueri ( Purkayastha et al. 2021) is ca. 78 km towards the south, in the State capital, Aizawl. The species is found to be nocturnal and all the specimens of C. vairengtensis sp. nov. were collected between ca. 2100-2240 h, from ca. 80 cm to 260 cm above ground. The holotype and two paratypes (MZMU2904 & MZMU2905) were on jointed weathered siltstone bed along a public link road ( Fig. 6A View FIGURE 6 ), whilst other paratypes were collected from a rocky wall covered by vegetation along public light vehicle road ( Fig. 6B View FIGURE 6 ) and stone steps ( Fig. 6C View FIGURE 6 ). All the specimens were collected from anthropogenic areas fringed with secondary forest in the peripheral part of the town. Climatic conditions in the area varied between 15°C and 21°C during winters and 27°C to 34°C during summers; the annual average rainfall is 2,674.5 mm ( Guhathakurta et al. 2020). The forest type of the location is tropical semievergreen forest as per the classification by Singh et al. (2002), corresponding to the semievergreen 2B/C2 forest type ( Champion & Seth 1968), and the vegetation around the area comprises mixed plantation trees such as Areca catechu, Artocarpus heterophyllus and Tectona grandis . Other common plants include Amomum dealbatum, Ananas comosus, Blumea lanceolaria, Manihot esculenta, Melocanna baccifera, Mikania micrantha, Musa paradisiaca, Mussaendra roxburghii and Thysanolaena maxima . Ferns such as Adiantum pedatum , A. aleuticum, Athyrium felix, Diplazium esculentum and Dryopteris filix were also abundant.
Etymology. The specific epithet is derived from the name of Vairengte town, Kolasib District of the state of Mizoram from where the type series were collected.
Suggested common name. Vairengte bent-toed gecko




| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |