
Pseudaeginella arraialensis, Ros, Macarena, Lacerda, Mariana B. & Guerra-García, José M., 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4258.4.6 |
|
publication LSID |
lsid:zoobank.org:pub:48DC505F-A448-4A6C-BCE7-0C0015B93326 |
|
DOI |
https://doi.org/10.5281/zenodo.6020848 |
|
persistent identifier |
https://treatment.plazi.org/id/039887C1-543C-986B-FF09-F92FFE96FDD0 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudaeginella arraialensis |
| status |
sp. nov. |
Pseudaeginella arraialensis sp. nov.
( Figs. 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Type material. Holotype, mature male (used for description and drawings of lateral view, antennae, gnathopods, pereopods 3–7 and abdomen) (vial and three slides, mouthparts dissected) (vial: MNCN 20.04 About MNCN /10341, slides: MNCN 20.04 About MNCN /10341a, MNCN 20.04 About MNCN /10341b, MNCN 20.04 About MNCN /10341c), Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 0.5–2 meters depth, associated to the hydroid Eudendrium sp., 16 October 2012. GoogleMaps
Paratype “a”, mature female (used for description and drawings of lateral view, gnathopod 2 and abdomen) (vial, mouthparts not dissected) (vial: MNCN 20.04/10342), collected together with the holotype.
Paratype “b”, mature male (vial and one slide, mouthparts dissected) (vial: MNCN 20.04/10343, slide: MNCN 20.04/10343a), collected together with the holotype.
Paratype “c”, mature male (vial and one slide, mouthparts dissected) (vial: MNCN MNCN 20.04/10344, slide: MNCN 20.04/10344a), collected together with the holotype.
Paratype “d”, mature male (vial and one slide, mouthparts dissected) (vial: MNCN 20.04/10345, slide: MNCN 20.04/10345a), collected together with the holotype.
Paratype “e”, mature female (vial and one slide, mouthparts dissected) (vial: MNCN 20.04/10346, slide: MNCN 20.04/10346a), collected together with the holotype.
Paratype “f”, mature female (vial and one slide, mouthparts dissected) (vial: MNCN 20.04/10347, slide: MNCN 20.04/10347a), collected together with the holotype.
Other paratypes (all collected together with the holotype): 3 mature males (non-dissected) (vials: MNCN 20.04/10348, MNCN 20.04/10349, MNCN 20.04/10350), 3 mature females (non-dissected) (vials: MNCN 20.04/ 10351, MNCN 20.04/10352, MNCN 20.04/10353). 5 mature males, 7 females (2 premature + 5 mature), 2 juveniles (non-dissected) (MNRJ 26601), 4 mature males, 6 females (2 premature + 4 mature) (non-dissected) (MCZ 134393).
Type locality. Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 0.5–2 meters depth. GoogleMaps
Additional material examined. 22 mature males, 28 females (21 premature + 17 mature), 18 juveniles, Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 0.5–2 meters depth, associated to the hydroid Eudendrium sp., 16 October 2012 GoogleMaps .
7 mature males, 4 females (2 premature + 2 mature), Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 0.5–2 meters depth, associated to the hydroid Ectopleura sp., 16 October 2012. GoogleMaps
2 mature males, 4 mature females, 2 juveniles, Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 0.5–2 meters depth, associated to the macroalgae Chylocladia sp., 16 October 2012. GoogleMaps
5 mature males, Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 3–5 meters depth, associated to the macroalgae Ceramium sp., 17 October 2012. GoogleMaps
2 mature males, 1 premature female Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 3–5 meters depth, associated to the macroalgae Chylocladia sp., 17 October 2012. GoogleMaps
1 mature male, Praia do Forno (22°57’58’’S, 42°00’27’’W), Arraial do Cabo, Rio de Janeiro, Brazil, 3–5 meters depth, associated to the macroalgae Corallina sp., 17 October 2012. GoogleMaps
Diagnosis. Head and pereonite 1 with a dorsal acute projection, bent forward. Pereonite 2 with a dorsal projection medially, a marked hump distally, and lateral acute projections (one in each side) near the coxa of gnathopod 2. Basis and ischium of gnathopod 2 with a distal projection. Antennae 1 and 2 setose in males. Gnathopod 2 very setose in males, provided with 1 middle-distal projection and 2 additional smaller distal projections.
Etymology. The specific name ‘ arraialensis ’ indicates the locality where the species was found, Arraial do Cabo.
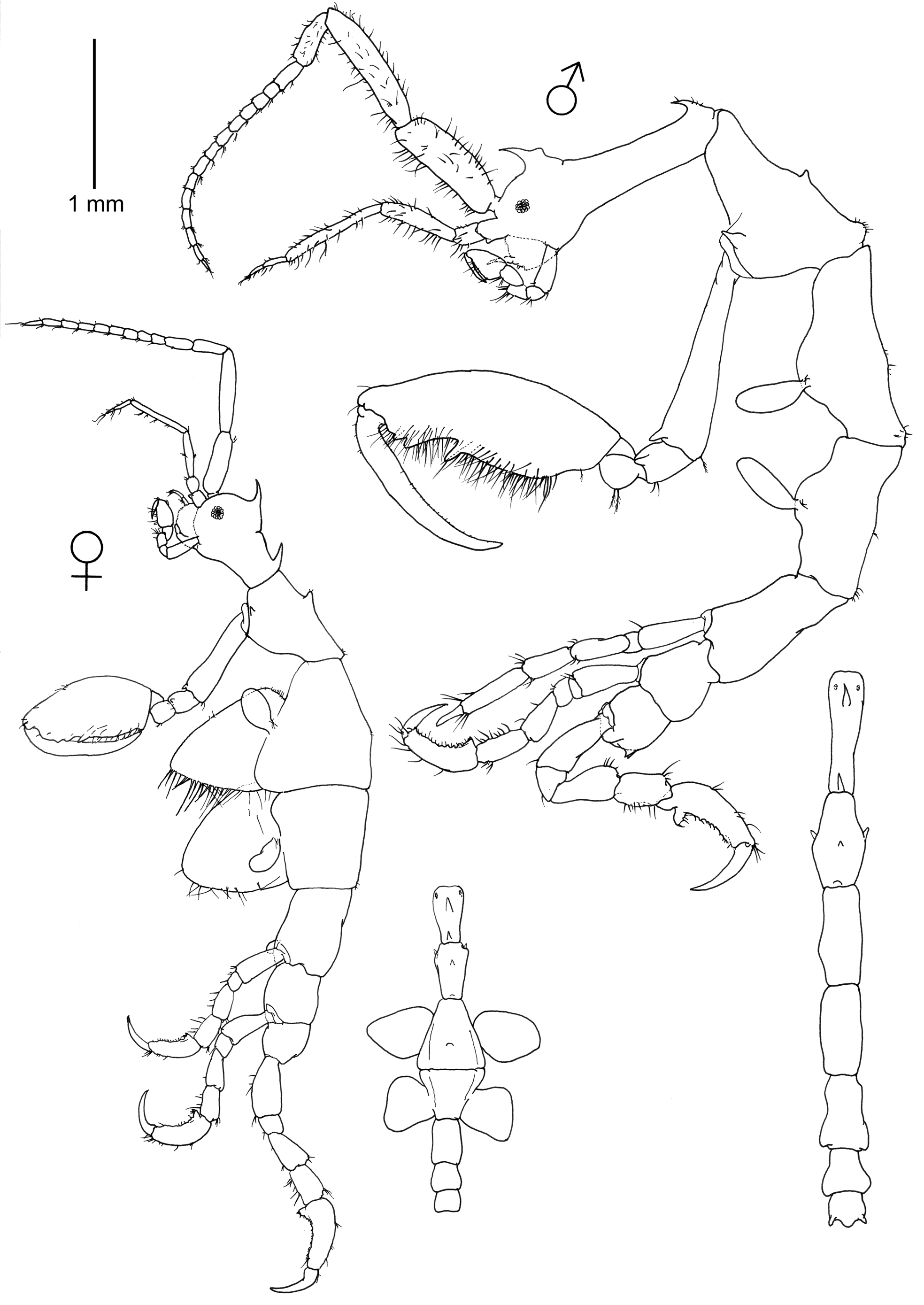
Description. Holotype male (6.7 mm) ( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Lateral and dorsal view ( Fig. 1 View FIGURE 1 ). Head with a dorsal acute projection, bent forward. The projection is provided with a dorso-medial rounded process. Eyes present and distinctive. Pereonite 1 fused with head, suture present. Pereonite 1 with another dorsal acute and forward bent projection distally. Pereonite 2 with a dorsal projection medially, a marked hump distally, and lateral acute projections (one in each side) near the coxa of gnathopod 2. Pereonites 3–6 without marked projections, only with medial and distal slight humps in pereonites 3 and 4. Pereonite 7 with two small dorsal projections laterally. Pereonite 3 the longest. Pereonite 7 the shortest.
Gills ( Fig. 1 View FIGURE 1 ). Present at middle of pereonites 3–4, elongate, length about 2.5 times width.
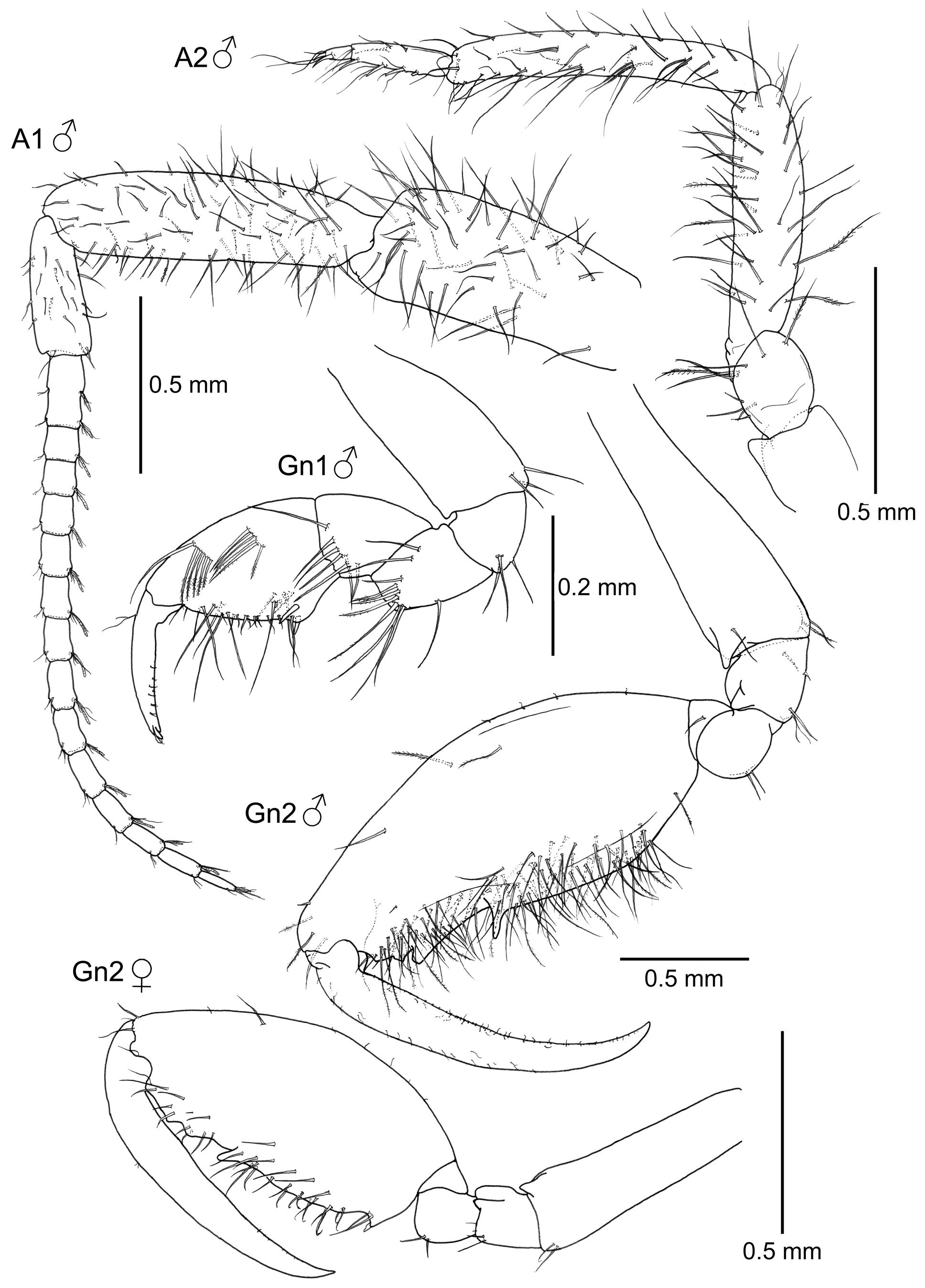
Mouthparts ( Fig. 2 View FIGURE 2 ). Upper lip symmetrically bilobed, smooth apically. Mandibles with no sign of mandibular molar. Three-articulate palp with a setal formula 1-x- 1, being x= 7 in left mandible and x= 8 in right mandible, medial article provided with three setae; left mandible with incisor 5-toothed, lacinia mobilis 5-toothed, followed by 2–3 accessory blades; right mandible with incisor 5-toothed, lacinia mobilis looking like a blade, followed by 2 more blades; no sign of molar flake. Lower lip without setulae; inner lobes rounded and almost fused. Maxilla 1 outer lobe carrying 6 serrated spines, palp 2-articulate, distal article with 3 apical spines and 1 medial seta. Maxilla 2 inner lobe small, shorter than outer lobe, with 4 and 5 setae respectively. Maxilliped inner plate small and rounded, about 1/3 of outer plate in length, carrying two apical setae; outer plate elongate, with 3 apical setae; palp 4-articulate, setose, distal article (dactylus) curved and provided with a proximal seta and rows of minute setulae.
Antennae ( Figs. 1 View FIGURE 1 and 3 View FIGURE 3 ). Antenna 1 peduncle with abundant setae; article 1 robust, second article the longest; third article the shortest; flagellum with 14 articles. First article composed of two articles fused. Antenna 2 setose, slightly shorter than peduncle of antenna 1; proximal peduncular article with a distal projection (gland cone); swimming setae absent; flagellum 2-articulate.
Gnathopods ( Figs. 1 View FIGURE 1 and 3 View FIGURE 3 ). Gnathopod 1 basis of the same length as the combination of ischium, merus and carpus; grasping margin of propodus non-serrated; 2 proximal grasping spines, dactylus with short setulae, bifid distally. Gnathopod 2 inserted on the posterior half of pereonite 2; basis longer than pereonite 2, with a distal acute projection; ischium small, rectangular with a distal acute projection; merus rounded; carpus short and triangular; propodus of gnathopod 2 elongate, very setose, 1 medio-distal projection and 2 additional smaller distal projections; dactylus with minute setulae.
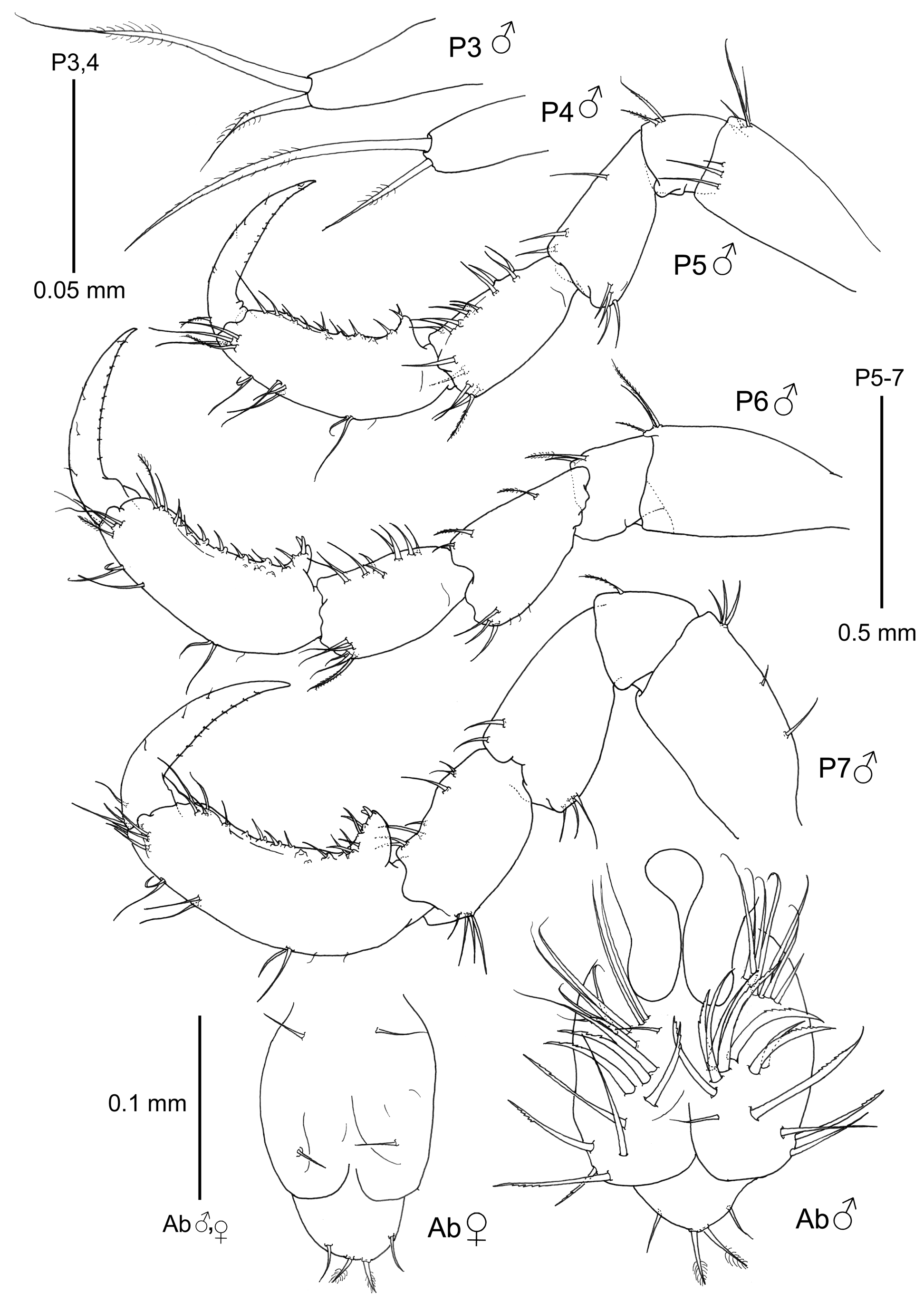
Pereopods ( Figs. 1 View FIGURE 1 and 4 View FIGURE 4 ). Pereopod 3 and 4 vestigial (ca. 0.05 mm), 1-articulate, provided with 2 distal plumose setae. Pereopod 5–7 similar in feature but increasing in robustness respectively; propodus with small tubercles on the inner margin; a proximal triangular projection provided with two grasping spines (in pereopod 5 these grasping spines are not so evident looking like setae); dactylus with row of fine setulae.
Penes ( Fig. 4 View FIGURE 4 ) large, elongate, length ca. 2.5 times width.
Abdomen ( Fig. 4 View FIGURE 4 ) without appendages, two lateral lobes with serrated setae and a single dorsal lobe with 2 plumose setae and 2 additional smooth setae
Paratype female “a” (4.2 mm) ( Figs. 1 View FIGURE 1 , 3 View FIGURE 3 , 4 View FIGURE 4 )
Antennae 1, 2 less setose than in male; flagellum of antenna 1 with 10 articles. Pereonite 1 less elongate than male. Pattern of dorsal projection as in male excepting for a marked hump medially in pereonite 3. Gnathopod 2 inserted on the anterior half of pereonite 2, propodus smaller than in male, less setose, and carrying a grasping spine proximally. Oostegites present on pereonites 3 and 4, the former being more setose than the latter. Pereonite 7 lacking lateral distal projections. Gills less elongate than in male. Lateral lobes of abdomen less setose than in male, provided only with two pairs of setae.
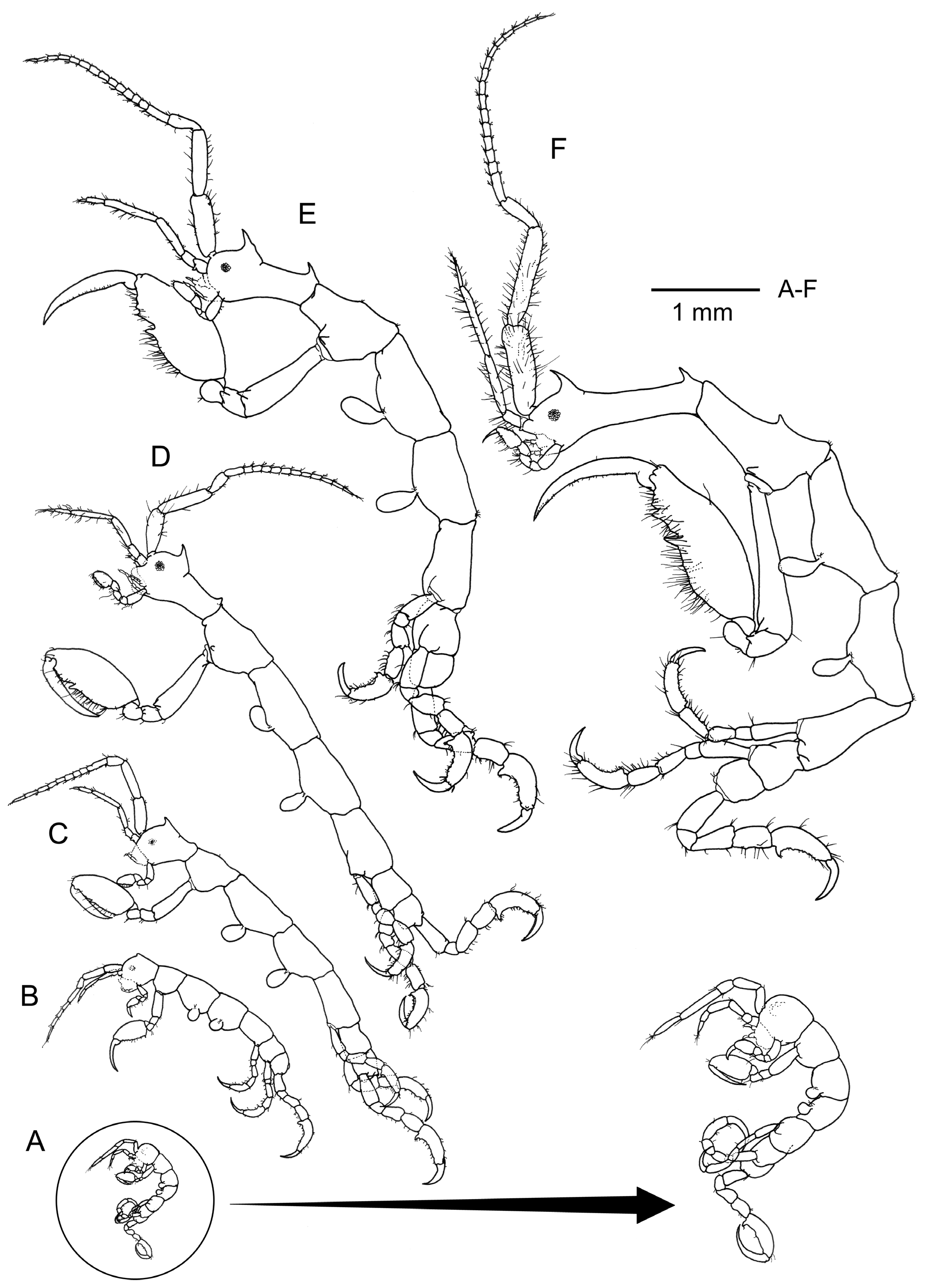
Intraspecific variation and ontogenetic development. The morphological characters of the species are rather constant in all the specimens examined. All mature males and females have a similar pattern of dorsal projections and humps: 1 acute projection on the head, 1 acute projection on pereonite 1, 1 projection medially and a marked hump distally on pereonite 2. Some mature females are provided also with a marked hump medially on pereonite 3. All mature specimens (both males and females) are also provided with a lateral projection near the coxa of gnathopod 2, and basis and ischium have also distal acute projections. A small round projection proximally on the basis of gnathopod 2 was observed in an adult male (see Fig. 5 View FIGURE 5 F), although this projection was absent in the holotype. The holotype has a pair of small projections dorso-distally on pereonite 7, which is absent in other males. Additionally, the dorsal projection of the head in the holotype is provided by a rounded process, which is absent in other males. The presence of this process, together with the lateral projections on pereonite 7, are probably characters only present in superadult males. The morphology of antennae, gnathopods and pereopods is similar in all mature specimens examined. The mouthparts were similar in all the specimens dissected. Inner and outer lobe of maxilliped were always provided with 2 and 3 apical setae respectively. Dactylus of the maxilliped palp always carries a single proximal seta, but the number of setae of articles 2 and 3 of the palp varies among specimens, increasing in number with the body size. All the mandibles examined were lacking molar process and were provided with 3 setae on article 2 of the palp. The setal formula was 1-x- 1 (with x being 7,8 or 9), except for two specimens, in which one of the mandibles was provided with 2 or 3 apical setae instead following the formula 1-x- 2 or 1-x- 3. Maxilla 1 was similar in all specimens, outer lobe provided with 6 setae and palp with 3+1 setae. Maxilla 2 was also similar in all the dissected specimens, with the outer lobe carrying 5 setae and inner lobe carrying 4 or 5 setae.
Regarding the ontogenetic development, newly hatched juveniles (instar 1, ca. 1.3 mm, see fig. 5) have no dorsal nor lateral projections, head and pereonite 1 are totally fused (suture not present), pereopods 3 and 4 are proportionally larger than in adult stages, flagellum on antenna 1 is 2-articulate, gills are rounded, eyes are not clearly distinguished, dactylus of pereopods is proportionally more developed than in adult stages. The first projection which can be distinguished is the projection of the head, which is already present in juveniles of 2 mm. Lateral projections near the coxa and distal projections in basis and ischium of the gnathopod 2 began to be present in juveniles of 3–4 mm. The number of setae in antennae and gnathopod 2 is increasing with body size and the pereonite 1 is also increasing in length in mature males. The number of articles in the flagellum of antenna 1 is increasing from 2 (in juveniles of instar 1) to 14–16 in larger specimens ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ). Gills are becoming more elongate with increasing body size and lateral expansions of pereonite 3 and 4 are more evident in the larger males. In females, the insertion of the basis of gnathopod 2 is always on the anterior half of pereonite 2, while in males is located on the anterior half in juvenile stages but on the posterior half in larger specimens.
Taxonomical remarks. Lacerda et al. (2011) provided a key to species of the genus Pseudaeginella and body illustrations of 9 known species of the genus. Presently, 11 species have been described: P. tristanensis ( Stebbing, 1888) from Tristan da Cunha, South Atlantic Ocean, P. antiguae Barnard, 1932 from Antigua (presently nomem dubium), P. biscaynensis (McCain, 1968) from Florida, USA, P. montoucheti from the coast of Brazil, P. polynesica (Müller, 1990) from Bora Bora and Moorea, French Polynesia, P. sanctipauli Laubitz, 1995 from St. Paul and Amsterdam Islands, P. campbellensis Guerra-García, 2003 from the subantarctic islands of New Zealand, P. vaderi Guerra-García, 2004 from Northern Territory, Australia, P. inae Krapp-Schickel & Guerra-García, 2005 from Indonesia, P. colombiensis Guerra-García, Krapp-Schickel & Müller, 2006 from the Caribbean coast of Colombia and P. hormozensis Momtazi & Sari, 2013 from the Gulf of Oman and the Persian Gulf.
The new species can be distinguished easily from the remaining species in the genus mainly by the pattern of dorsal projections, and by the setose antennae and gnathopod 2 in males. The closest species to P. arraialensis seems to be P. tristanensis (see Stebbing, 1888; Laubitz, 1995). This species was initially described by Stebbing (1888) as Aeginella tristanensis based on a single immature specimen collected from Tristan da Cunha. The length of this species was reported as “one-seventh of an inch”, which correspond to 3.63 mm. When we compare the figures of this immature specimen with the equivalent immature of P. arraialensis (see Fig. 5 View FIGURE 5 C), a clear difference in the shape of the head projection can be observed: immature of P. tristanensis is provided with a small, straight and rounded projection while P. arraialensis juveniles have a bent forward acute projection. Furthermore, the pereonites 3 and 4 are clearly more elongate in the juvenile of P. tristanensis than in P. arraialensis . After the description of Stebbing (1888), Mayer (1890) erected the new genus Pseudaeginella and transferred the species to this new genus, providing figures of abdomen and maxilliped. Laubitz (1995) collected material from Ile Amsterdam and identified this as P. tristanensis . She pointed out that it is clear that changes in proportions of body and gnathopods occur when the male reaches maturity. The largest male studied by Laubitz (1995) was 6.5 mm, the same order than male specimens of P. arraialensis in the present study. The most relevant differences between P. arraialensis and P. tristanensis of Laubitz (1995) are: (1) pereonite 4 is provided with a clear dorsal hump distally in P. tristanensis which is lacking in P. arraialensis , (2) pereonite 5–7 have small acute projections in P. tristanensis while in P. arraialensis the projections are only present in pereonite 7 of some males, (3) antennae and gnathopod 2 are much more setose in males of P. arraialensis , (4) setal formula (1-x- 1) of mandible palp with x= 7– 9 in P. arraialensis and x= 5 in P. tristanensis .
The number of dorsal body projections is usually increasing with age of specimens. The male of P. tristanensis figured by Laubitz (1995: 89, fig. 4C) is apparently less developed than the adult males of P. arraialensis of the present study ( Fig. 1 View FIGURE 1 , Fig 5 View FIGURE 5 F) since it has the pereopod 2 less elongate and the insertion of gnathopod 2 basis is located on the anterior end of pereonite instead of the posterior end. In spite of this, the number of dorsal projections and humps is higher in this male of P. tristanensis , with dorsal hump in pereonite 4 and acute projections in pereonites 5–7. Pereonites 4, 5 and 6 are totally smooth in all specimens examined of P. arraialensis .
During a review of caprellids from the Red Sea, Zeina & Guerra-García (2016) suggested the genus Paradeutella could be considered as synonymous of Pseudaeginella . Both genera were described in the same year ( Mayer, 1890). Several species of Pseudaeginella have been recently described and most of them include detailed taxonomic description according to the modern taxonomy, so, according to Zeina & Guerra-García (2016) it would be advisable to maintain Pseudaeginella as valid genus and Paradeutella as synonymous. Most of the Paradeutella species are based on very poor and incomplete descriptions, and need redescriptions. Pseudaeginella arraialensis is close to Paradeutella bidentata from India, although (1) P. bidentata lacks the dorsal acute projection on pereonite 1, (2) propodus of gnathopod 2 male is elongate in P. arraialensis and lacks medial projection, while it is oval in P. bidentata and it is provided with a proximal grasping spine and medial projection and (3) male antennae and gnathopod 2 are clearly more setose in P. arraialensis .
Dietary analysis. Gut contents of the 24 analysed specimens included detritus, microalgae and crustaceans. The component which occupied the majority of the gut area was detritus (45 %) followed by rest of crustaceans (31%) and microalgae (24%). This indicates that Pseudaeginella arraialensis can be considered as omnivorous species. A relative high percentage of crustaceans, together with a considerable number of empty guts found, suggest the importance of carnivory in this species. These observations are supported by the lack of molar in this species, since Guerra-García et al. (2014) found that in carnivorous taxa the mandibular molar is usually lacking or very reduced, while detritivorous species are characterised by a stronger molar.
In general terms caprellids feed mainly on detritus, and many species are opportunistic (Guerra-García & Tierno de Figueroa, 2009; Guerra-García et al., 2014). The only Pseudaeginella species studied regarding the diet are P. colombiensis and P. inae , characterised as detritivorous (Guerra-García & Tierno de Figueroa, 2009), although the number of specimens examined was very low.
Taking into account that the highest abundances of P. arraialensis were found in the hydroid Eudendrium sp, a cleptocommensalist behaviour might occur by stealing captured prey from the polyps of the hydroid. This trophic strategy was observed by Bavestrello et al. (1996) for Pseudoprotella phasma , and Ros & Guerra-García (2012) for Paracaprella pusilla .
Habitat use patterns. The species exhibited a high plasticity to inhabit different host substrata and habitat types, since it was found attached to hydroids ( Eudendrium sp. and Ectopleura sp.) and macroalgae ( Corallina sp. and Chylocladia sp.), both in fouling communities of artificial floating structures (floating restaurant and oyster culture) and nearby natural habitat (rocky shore). However, it is worth noting that the species was not found in a previous study conducted by Serejo (1998) regarding the amphipod fauna associated with the sponge Dysidea fragilis Johnston at Arraial do Cabo. Although P. arraialensis did not show a specific relationship with any substratum in particular, the highest densities were found attached to hydroids of artificial substrata. This indicates that the species could be prone to live in this kind of artificial habitats and, therefore, to anthropogenic dispersion (e.g. through the importation of living Pacific oysters). However, this possibility should be taken with caution since not all species inhabiting artificial habitats are anthropogenically dispersed. In fact, Ros et al. (2016) found two caprellid species endemic to Brazil ( P. montoucheti and Aciconula tridentata Guedes-Silva and Souza-Filho, 2013 ) also inhabiting artificial habitats (marinas). Interestingly, P. arraialensis has not been found yet in other areas of southern Brazil in spite of the extensive collections conducted with a similar methodology (see Ros et al, 2016). Although the distribution of this species seems to be very restricted, its abundance in the type locality is high. A similar situation has been reported also for other caprellids; it is the case, for example, of Deutella venenosa Mayer, 1890 . This species is locally very abundant in the fouling communities of buoys in Coquimbo, Chile, being one of the dominant species ( Guerra-García & Thiel, 2001). However the species shows a very restricted distribution since it has been only found in Coquimbo, being absent in other areas of Northern-central Chile ( Thiel et al., 2003). It is possible that the special environmental conditions occurring at Arraial do Cabo could contribute to the restricted distribution of P. arraialensis . Further studies in other different areas of Brazil are still needed to confirm this possibility.


| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Caprellinae |
|
Genus |