
Copionodon exotatos, Pinna & Abrahão & Reis & Zanata, 2018
|
publication ID |
https://doi.org/ 10.1590/1982-0224-20180049 |
|
publication LSID |
lsid:zoobank.org:pub:661304D3-1725-402C-B7E4-E7DD13009864 |
|
DOI |
https://doi.org/10.5281/zenodo.3717000 |
|
persistent identifier |
https://treatment.plazi.org/id/2D56692E-B1F3-4557-89C6-0595CCE4CA9C |
|
taxon LSID |
lsid:zoobank.org:act:2D56692E-B1F3-4557-89C6-0595CCE4CA9C |
|
treatment provided by |
Carolina |
|
scientific name |
Copionodon exotatos |
| status |
sp. nov. |
Copionodon exotatos , new species
urn:lsid:zoobank.org:act:2D56692E-B1F3-4557-89C6-0595CCE4CA9C
( Figs. 1-3 View Fig View Fig View Fig )
Holotype. MZUSP 123522 View Materials , 36.9 mm SL, Brazil, Bahia, Chapada Diamantina, Lençóis , exit of rock-enclosed sector of right-hand branch of riacho do Mosquito (immediately upstream from Cachoeira do Mosquito ), itself a tributary of rio Santo Antônio , rio Paraguaçu basin 12°22’15.64”S, 41°22’15.85”W, M. de Pinna, V. Abrahão, V. Reis & A. Zanata, 4 Mar 2017. GoogleMaps
Paratypes. MZUSP 121656, 5, 12.6-35.8 mm SL, collected with holotype GoogleMaps .
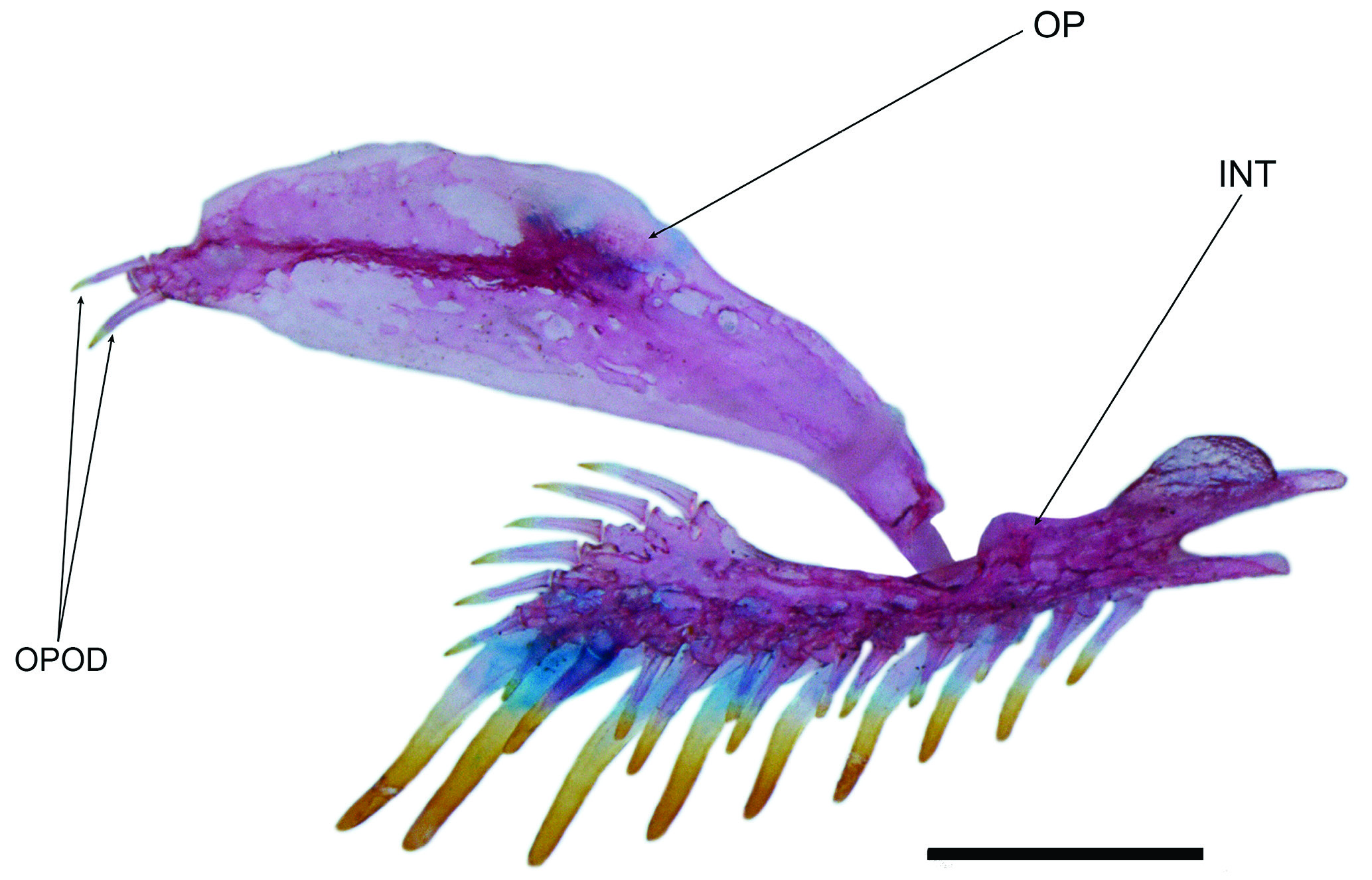
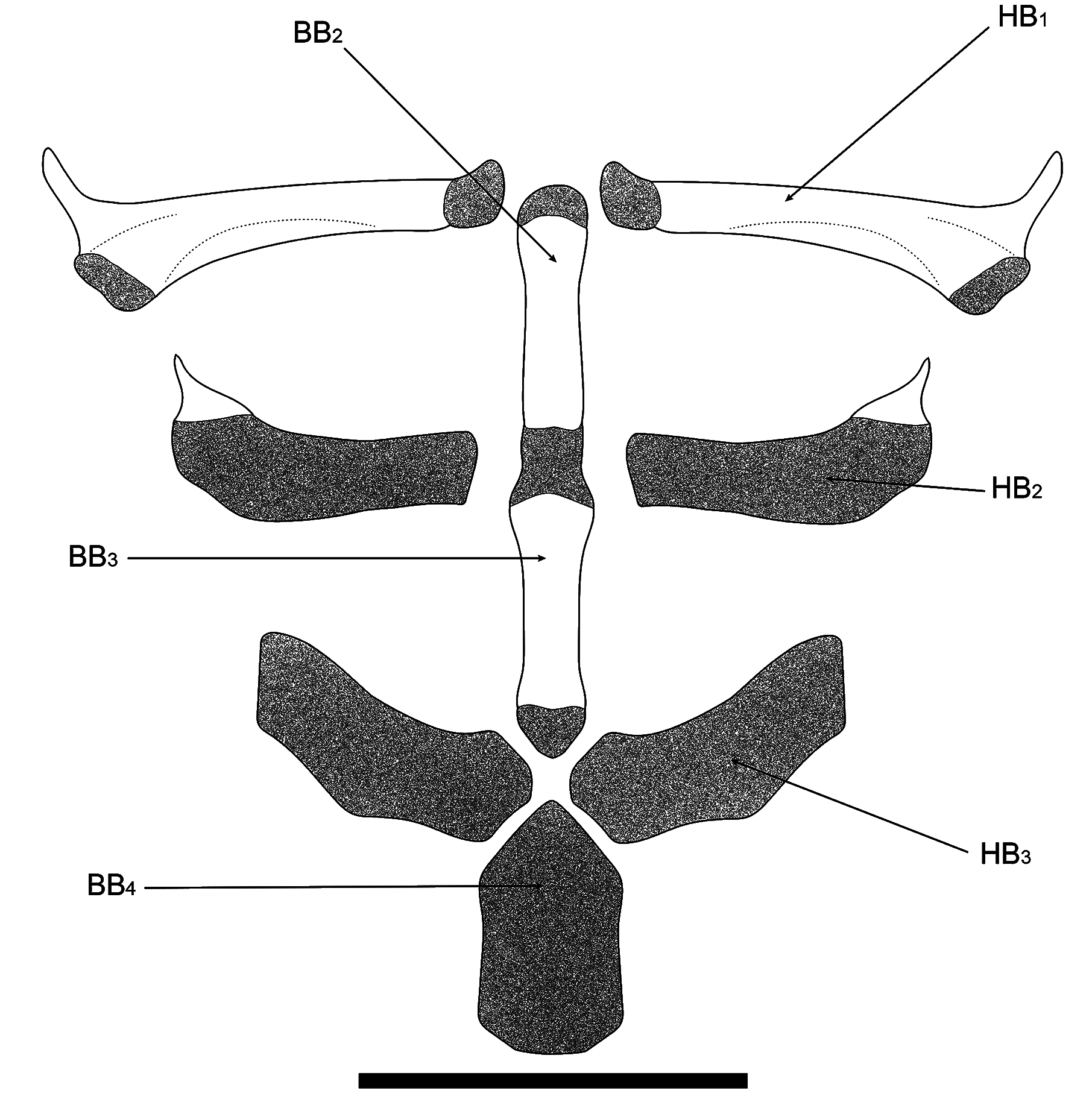
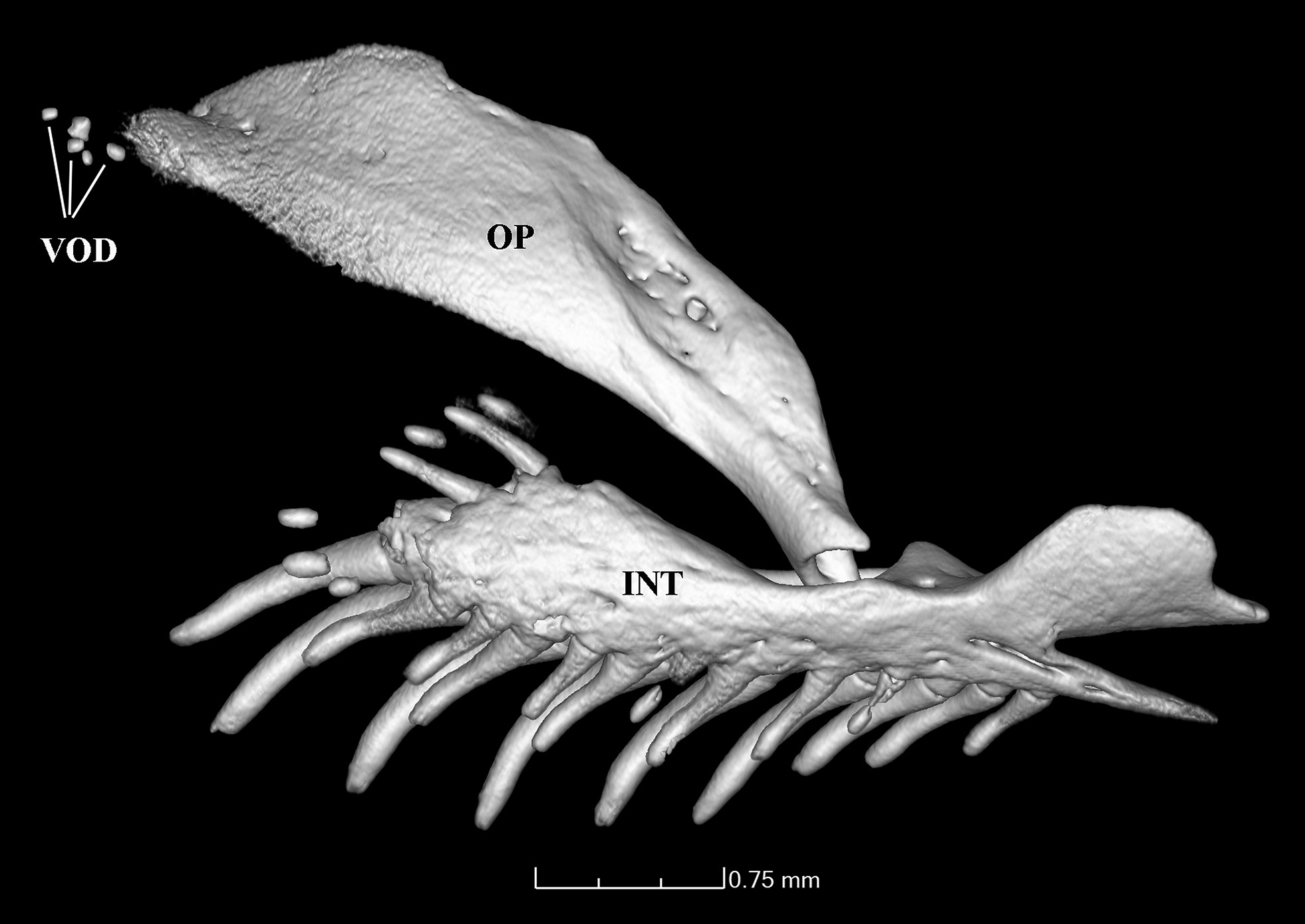
Diagnosis. Distinguished from all copionodontines, except Glaphyropoma spinosum , by the presence of opercular odontodes (with presence variable; vs. opercular odontodes entirely absent; Figs. 2-4 View Fig View Fig View Fig ). Further distinguished from all congeners by the small eye in mid- to large-sized specimens (over 35 mm SL, 11-12% HL; vs. 13% or more; Fig. 2a View Fig ); and from all except C. elysium de Pinna, Burger & Zanata, 2018 by the lack of a free orbital rim ( Fig. 2a View Fig ). The combination of body coloration lacking conspicuous markings and lack of free orbital rim distinguishes C. exotatos from all other congeners. Distinguished from C. elysium , C. pecten de Pinna, 1992 and C. orthiocarinatus de Pinna, 1992 by the color pattern composed of a tenuous continuous and straight midlateral stripe against a uniform scattering of dark chromatophores ( Fig. 1 View Fig ; vs. color pattern more complex, including irregular dark markings dorsal and ventral to an irregular longitudinal stripe in C. elysium and C. pecten or absence of a midlateral stripe in C. orthiocarinatus ). Further distinguished from C. elysium by the presence of a conical ossification cap in the anterolateral corner of the second hypobranchial ( Fig. 5 View Fig ; vs. second hypobranchial entirely cartilaginous; this ossification is so far verified in a single c&s paratype of C. exotatos ). Copionodon exotatos further differs from C. pecten by the smaller interopercular patch of odontodes, its anterior limit distant from lower lip (by distance equivalent to approximately mouth width) and posteriorly not reaching base of first ral-fin origin and rictal one extending beyond midlength of interopercular patch of odontodes ( Figs. 1-2 View Fig View Fig ; vs. maxillary barbel reaching posterior ¼ of length of interopercular patch of odontodes and rictal reaching anterior ¼ of length of interopercular patch of odontodes). From species of Glaphyropoma , the new taxon further differs by the broad first hypo- branchial ( Fig. 5 View Fig ; vs. slender; cf. de Pinna, 1992, fig. 6 and Bichuette et al., 2008, fig. 6), the caudal fin bilobed ( Fig. 1 View Fig ; vs. truncate), the toothed portion on dentary not reaching the coronoid process ( Fig. 6 View Fig ; vs. reaching coronoid process), and the presence of asymmetrical spatulate teeth in jaws ( Fig. 11 View Fig ; vs. all teeth symmetrically spatulate).
pectoral-fin ray ( Fig. 2b View Fig ; vs. interopercular patch of odontodes large, its anterior limit near lower lip (separated by distance equivalent approximately to half of mouth width) and posteriorly reaching beyond pectoral-fin base) and lon- ger barbels, with maxillary barbel usually reaching pecto- Description. Body elongate, trunk roughly as deep as wide in cross-section near head, gradually more compressed posterior to end of pectoral fin. Caudal peduncle compressed, tapering to caudal fin. Dorsal profile of body and head ne- arly straight from snout to dorsal-fin origin, slightly convex near head. Posterior region of body straight from endpoint of dorsal fin to caudal-fin origin. Ventral profile straight along head and gular regions, nearly parallel to longitudinal axis of body; convex or slightly convex from that point to pelvic-fin origin and nearly straight or slightly concave from pelvic-fin base to caudal-fin base. Dorsal profile of adipose fin nearly straight; fin barely developed, somewhat more visible pos- terior to vertical through end of anal-fin base. Adipose and caudal fins confluent ( Figs. 1-3 View Fig View Fig View Fig ).
Head slightly longer than wide and depressed. Mouth inferior, subterminal, located close to anterior margin of snout. Upper lip narrow in ventral view, rounded ventrally. Ventral margin of upper lip continuous laterally with rictal barbel. Lower lip with about same thickness as upper lip; continuous, not divided in lateral halves by median constriction, only slightly less convex. Angle of mouth with fleshy outgrowth just posterior and adpressed to rictal barbel ( Figs. 2-3 View Fig View Fig ).
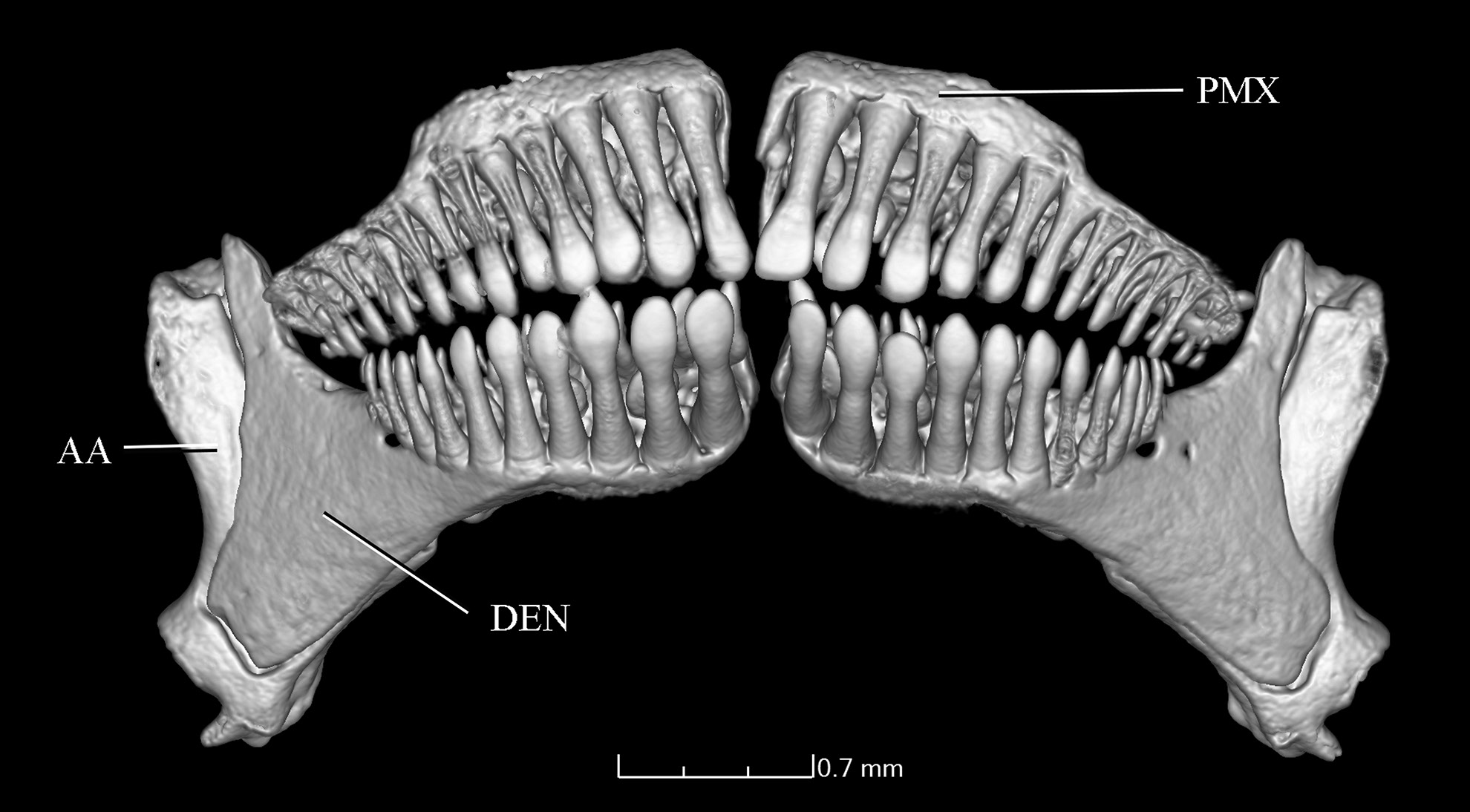
Dentary and premaxillary teeth somewhat similar to each other in shape and arrangement ( Fig. 11 View Fig ). Teeth gradually larger towards symphysis. First row of teeth spatulate and darkly-pigmented distally in dentary and premaxilla, with spatulate portion in some teeth symmetrical and others asymmetrical; spatulate portion not overlapping in lower jaw ( Fig. 11 View Fig ). Distal border of teeth straight. In both jaws, second row of teeth also slightly expanded distally; third row conical. Teeth on lower jaw distributed over anterior two thirds of ossification and not reaching coronoid process. Outer series of lower jaw with about six to eight spatulate larger anterior teeth followed by three to five conical teeth.
Eyes large in small specimens but small in mid- to large- -sized ones, without free orbital rim and covered with transparent skin. Eyes located slightly anterior to half of head length, much closer to lateral border of head than to midline in dorsal view. Infraorbital latero-sensory canal complete, extending from sphenotic posteriorly to first infraorbital (la- crimal) anteriorly. Three infraorbital pores, one directly posterior to posterior margin of eye, another directly ventral to ventralmost point of eye, and a third anteroventral to eye in lateral view. Anterior nares surrounded by short tube of integument directed anterolaterally, continuous posterolaterally with nasal barbel. Posterior nares slightly closer together than anterior ones, surrounded by tube of integument incomple- te posterolaterally, with appearance of two partly confluent flaps of skin. Three pairs of thin flat barbels, wide at base and gradually narrowing distally to fine tip. Maxillary barbel reaching slightly beyond base of pectoral fin. Rictal barbel inserted ventral to maxillary barbel, extending to middle of interopercular patch of odontodes. Nasal barbel originating on posterolateral region of anterior nares, reaching to middle of opercular patch of odontodes (or homologous position in specimens without latter) ( Figs. 2-3 View Fig View Fig ). Nasal barbels shorter in juvenile specimens (12.6 and 13.1 mm SL), reaching to middle or posterior margin of eye. Interopercle with well- -developed odontodes, visible in ventral aspect of head. Interopercular patch of odontodes extending from vertical through posterior border of eye anteriorly to slightly anterior to vertical through pectoral-fin base posteriorly. Odontodes arranged in two series, with those on mesial series larger than those on lateral one; odontodes gradually larger posteriorly in both series, with those posteriorly on mesial row largest. Eight odontodes on mesial row and six or seven on lateral row. Interopercular odontodes not yet developed in two smallest specimens examined (12.6 and 13.1 mm SL). Opercular patch of odontodes variably present in two of four adults or subadult type specimens (odontodes not yet developed in two additional juvenile paratypes). Opercular odontodes vestigial in holotype, apparently lacking direct mineralized connection with opercle ( Fig. 13 View Fig ) yet visible on surface of skin. Patch small and inconspicuous on surface of head, located anterodorsally to pectoral-fin base, with single row of four or five small odontodes sunk in individual slits of integument. Entire patch surrounded by specialized rim of integument typical of that complex in most other trichomycterids. Specimens without opercular patch of odontodes with homologous region of head smooth, lacking both odontodes and associated integumentary modifications ( Fig. 4 View Fig ).
Pectoral fin large, extending to middle of distance betwe- en occipital and pelvic-fin base, gently convex in distal pro- file, its base immediately posterior to vertical through pos- terior tip of interopercle ( Fig. 1 View Fig ). Pectoral-fin rays i8i*(1), i9(1), or i9i(2). Pectoral-fin rays approximately same length except for short last one. Pelvic fin with round distal pro- file, its origin at or slightly posterior to middle of SL, and well anterior to vertical through dorsal-fin origin, entirely covering anal and urogenital openings, reaching or almost reaching anal-fin origin ( Fig. 1 View Fig ). Pelvic-fin rays i5/i5i*(1) or i5i(3), first ray (unbranched) shorter than others. Dorsal fin smaller than anal fin, its distal profile gently convex. Dorsal-fin origin closer to base of caudal fin than to tip of snout ( Fig. 1 View Fig ). Dorsal-fin rays i5i*(1), ii5(3). Anal fin with base longer than that of dorsal, and total area larger than those of all other fins except caudal, its distal profile strongly convex. Anal-fin origin slightly anterior to vertical through end of dorsal-fin base ( Fig. 1 View Fig ). Anal-fin rays ii8*(1), iii9(1), or iii10(2). Caudal fin bilobed with round corners, with 8/9 principal rays. Upper lobe slightly longer than lower lobe in some specimens ( Fig. 1 View Fig ). Six dorsal and ventral procurrent caudal-fin rays, plus one dorsal segmented non-principal ray dorsally and two ventrally. Adipose fin shallow, without clear anterior and posterior limits, extending between end of dorsal fin and caudal-fin origin ( Fig. 1 View Fig ). Degree of deve- lopment of adipose fin ontogenetically variable, apparently inversely proportional do size of fish. Vertebrae 33*(2) or 34(2). First dorsal-fin pterygiophore immediately posterior to neural spine of 11th or 12th vertebra, first anal-fin ptery- giophore immediately posterior to neural spine of 14th or 15th vertebra.
Lateral line splitting immediately after leaving neurocranium, with ventral branch very short and with single terminal pore. Dorsal and main branch extending continuously to approximately a vertical through middle of pectoral fin and extending posteriorly along entire body length as series of independent tubules, progressively shorter and more spaced out posteriorly. Last tubule over complex caudal centrum and base of hypural plate. Lateral line ossicles absent. Pleural ribs 11*(4). Branchiostegal rays six* (4). Dorsal-fin pterygiophores seven* (4). Anal-fin pterygiophores 12(2); remaining specimens could not be examined with the available images. A single large and ossified proximal pectoral-fin radial and four (1) or six (1) cartilaginous distal pectoral-fin radials.
Coloration in alcohol. Dorsum and sides of body mostly uniformly colored by small dark chromatophores, dense dorsally ( Figs. 1-3 View Fig View Fig View Fig ). In holotype, faint lateral dark band extending midlaterally alongside of body, more pronounced
posterior to vertical through middle of dorsal-fin base. Band running exactly along midlateral line on the anterior half of SL, shifting to immediately dorsal to midline posteriorly to that. Middorsal line slightly lighter than rest of dorsum anteriorly to dorsal fin in some specimens. Abdominal region mostly white, with sparse dark fields mesial to base of pec- toral fin ( Fig. 2b View Fig ).
Overall color of head similar to, and slightly denser than, that of dorsum. Region of head corresponding to skull roof darkest due to combination with underlying brain pigment. Laterosensory pores on head white. Nostrils whitish. Integument slits of individual opercular odontodes, when present, white. Ventral surface of head with scattered minute melanophores ( Fig. 2b View Fig ). Fleshy outgrowth at angle of mouth white. Upper and lower lips heavily darkly-pigmented, except for white anterior margin of lower lip and inner margin of upper one. Nasal barbel uniformly dark along its entire length, on both sides. Maxillary barbel lighter than nasal barbel and gradually lighter distally, its dorsal surface darker than ventral one. Rictal barbel mostly white, with few dark fields near base. All rayed fins with sparse melanophores irregu- larly scattered alongside individual rays and segment limits. Pelvic and anal fins lighter than remaining fins. Adipose fin with uniform coating of melanophores similar to that on dorsal portion of body but looking darker because of different underlying tissues. Base of pectoral fin with rounded dark fields. Region around base of pelvic fins white. Narrow vertical white area at base of caudal fin, clearly separating origin of fin from caudal peduncle ( Figs 1-3 View Fig View Fig View Fig ).
Coloration of very small specimens (12.6 and 13.1 mm SL) similar to that of adults, but with a mostly white adipose fin and lower caudal peduncle fin-fold and no trace of a midlateral stripe.
Etymology. The specific epithet comes from the Greek exotatos , meaning outermost, in reference to the outlying locality of the new species.
Distribution. Known exclusively from the type-locality ( Fig. 7 View Fig ).
Ecological notes. The riacho do Mosquito is perennial and torrential in the rainy season, but its flow is reduced drasti- cally in the dry season. It runs on the eastern slopes of the Sincorá Range. The hydrological route of the stream at the type locality is peculiar. At approximately 12º21’59.51”S 41º22’19.37”W it splits into right and left arms. The entire right arm enters a sinkhole shortly after the split and becomes totally subterranean for ca. 500 m (in straight line, likely more as stream path; this subterranean sector is yet unmapped). The left arm remains on the surface. At the time of our sampling the entire region was undergoing the hardest drought in many years and the left epigean arm was entirely dry while the right arm maintained a small but consistent flow. The course of the two arms rejoin shortly after the emergence of the right arm, immediately before the precipitous waterfall called Cachoeira do Mosquito. Copionodon exotatos was sampled at a point near the emergence of the subterranean sector of the right arm ( Fig. 8 View Fig ). That portion is the terminal part of the subterranean sector and is itself not exactly a cave, but rather a rock-enclosed portion at ground level on sloped terrain, with part of the enclosure apparently man-made and with evidence of past use of explosives. Such structures are locally known as “grunas” and are sites of past small-scale diamond mining. All specimens came from a small area some 10 cm deep, but which was the deepest portion of the stream in several square meters. The substrate was gravel, sand and solid rock. Collection and visual inspection of the riacho do Mosquito about three km both upstream and downstream from the subterranean sector revealed no copionodontines, but its uppermost sectors remain unsampled. Examination of stomach contents of one paratype (now c&s) of C. exotatos includes allochthonous and autochthonous items, composed mainly of aquatic insects.
Conservation status. Copionodon exotatos is known exclusively from a short (ca. 500m) and mostly underground sector of the riacho do Mosquito (tributary of the Upper rio Paraguaçu basin, within the Chapada Diamantina Domain). It is possible that during the rainy season it spreads further out in the epigean sector, but this is currently unknown. The reduced flow of water in the dry season makes that stream particularly vulnerable to any pollutants and is associated with the entrenched distribution of the species, which in turn results in an obviously vulnerable situation. The environment of the species has well oxygenated fast-running clear water running over nude rocky bottom with patches of sand. The apparently narrowly endemic nature of the species makes it (and remaining copionodontines) dependent on the preservation the delicate headwaters they inhabit. The reduced flow of water in the dry season, and the use of water for human consumption and livestock in addition to deforestation of the banks constitute threats to the species, resulting in an obviously vulnerable situation. This, along with the absence of studies on population biology and geographical range reductions for this species allow us to classify C. exotatos as Data Deficient (DD) according to the International Union for Conservation of Nature (IUCN) categories and criteria (IUCN Standards and Petitions Subcommittee, 2017).
Remarks. Because no specimens of C. exotatos were found either upstream or downstream of the subterranean sector of the riacho do Mosquito, it is likely that most of the population of the species resides inside the subterranean portion, yet unsampled. In that case, the type specimens of C. exotatos were probably rogue specimens roaming close to the limits of their subterranean habitat, at the beginning of the outflowing epigean sector. Collection of more abundant ma- terial of the species, from throughout its range, will require speleological exploration of the Mosquito Cave.






| V |
Royal British Columbia Museum - Herbarium |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |