
Magelona guineensis, Mortimer & Kongsrud & Willassen, 2022
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlab070 |
|
publication LSID |
lsid:zoobank.org:pub:278AA1B0-674E-414D-A47A-D87F43E2D6E4 |
|
DOI |
https://doi.org/10.5281/zenodo.6459422 |
|
persistent identifier |
https://treatment.plazi.org/id/039087DB-FFED-FFF1-4245-742AFAD9FC0C |
|
treatment provided by |
Plazi (2022-04-13 07:05:41, last updated 2024-11-27 10:02:10) |
|
scientific name |
Magelona guineensis |
| status |
sp. nov. |
MAGELONA GUINEENSIS View in CoL SP. NOV.
( FIGS 7 View Figure 7 , 8 View Figure 8 )
Z o o b a n k r e g i s t r a t i o n. u r n: l s i d: z o o b a n k. org:act: 053593E9-5402-4D0F-9519-8CAC8C23788E.
Type locality: São Tomé and Príncipe, 1.5345°N 7.4225°E, 38 m depth GoogleMaps .
Type material: Holotype, São Tomé and Príncipe St. 5SP–05, af in 75% Etoh ( ZMBN132137 View Materials ) . Paratypes: Guinea (Conakry): St. 7GU–01, 5af in 75% Etoh ( ZMBN107270 View Materials ) ; 1af in 75% Etoh ( ZMBN107271 View Materials , imaged) . Sierra Leone: St. 7SL–04, 1af in 75% Etoh ( NMW.Z.2021.001.0004) . Nigeria: St. 6N–01, 1af in 96% Etoh ( ZMBN115736 View Materials , DNA-voucher); St. 6N–20, 1af in 96%Etoh ( ZMBN107331 View Materials , DNA-voucher) . Cameroon: St. 5C–10, 1af in 96% Etoh ( ZMBN107269 View Materials ) ; St. 6C–21, 3af in 75% Etoh ( ZMBN107268 View Materials ) . Gabon: St. 5G–13, 1af in 75% Etoh ( ZMBN132140 View Materials ) . São Tomé and Príncipe: St. 5SP –05, 1af, 1f in 75% Etoh ( NMW.Z.2021.001.0005); 1af in 96% Etoh ( ZMBN107303 View Materials , DNA-voucher) . Angola: St. 7AN–05, 1af in 75% Etoh ( ZMBN107267 View Materials ) .
Etymology: The specific name refers to type locality in the Gulf of Guinea.
Additional material: Guinea-Bissau: St. 6 GB – 07, 17af in 75%Etoh (ZMBN107272); St. 7 GB –08: 3af in 96%Etoh (ZMBN107273); 1af in 96%Etoh (ZMBN107274).
Diagnosis: Prostomium width similar to length, no prostomial horns. Chaetigers 1–8 with slender sinuous notopodial lamellae with small superior dorsal lobes and triangular ventral neuropodial lamellae. Lamellae of chaetiger 9 triangular and postchaetal, with additional small, ventral, neuropodial processes. All thoracic chaetae capillary. Abdominal lateral lamellae spatulate, with pointed tips. Abdominal hooks tridentate, in two groups, vis-à-vis. No pouches observed, pygidium unknown.
Description: A moderately sized species; junction between thorax and abdomen fairly marked ( Figs 7A View Figure 7 , 8A, B View Figure 8 ), abdomen more rounded than the dorsally flattened thorax. Holotype, anterior fragment: prostomium 0.7 mm wide, 0.7 mm long; thorax 3.5 mm long (including prostomium), 0.65 mm wide; abdomen, 0.45 mm wide; total length 9 mm for 21 chaetigers. Largest DNAvoucher specimen (ZMBN107331), anterior fragment: prostomium 0.6 mm long, 0.6 mm wide; thorax 5.0 mm long (including prostomium), 0.6 mm wide; abdomen 0.65 mm wide; total length approximately 7.8 mm for 17 chaetigers (width measurements not including parapodia). Other anterior fragments: 3–14 mm long for eight to 31 chaetigers.
Prostomium ( Figs 7B View Figure 7 , 8A, C View Figure 8 ), approximately as wide as long (L: W ratio 1), anterior margin straight and square, rounded lateral margins. One pair of prominent longitudinal dorsal muscular prostomial ridges, abutting for majority of length, diverging at distal tips. Thin and marginally shorter outer pair of ridges abutting inner pair for entire length. Light, almost diagonal lines, either side of ridges, composed of small oblong to long rectangular patches, not as marked as seen in other species. Burrowing organ partially everted in one specimen (ZMBN107331), longitudinally ridged. No palps retained.
Achaetous region behind the prostomium, roughly twice the size of chaetiger 1 ( Figs 7A View Figure 7 , 8A, C View Figure 8 ). Chaetigers 1–8 similar; parapodia biramous ( Fig. 7C–L View Figure 7 ). Notopodia with low, triangular prechaetal lamellae confluent with slender, smooth-edged, sinuous postchaetal lamellae, decreasing slightly in size along thorax. Single, small digitiform, prechaetal superior dorsal lobe present on all thoracic chaetigers, except chaetiger 9, decreasing in size in posterior thorax. Neuropodia with low pre- and postchaetal lamellae, encircling chaetae cufflike and confluent with slender triangular lamellae underneath the chaetal bundle, with rounded tips. Lamellae initially slightly prechaetal in position but becoming completely ventral by chaetiger 4, decreasing in size in posterior thorax.
Chaetiger 9: shorter and narrower than preceding chaetigers ( Figs 7A View Figure 7 , 8A, C View Figure 8 ). Notopodial prechaetal lamellae low, confluent with slender triangular postchaetal lamellae, shorter but wider than on preceding chaetiger ( Fig. 7M View Figure 7 ). No superior dorsal lobes observed. Neuropodia similar to notopodia, with a low inferior ridge terminating in an additional small, triangular process on both sides of the chaetiger. Chaetae of chaetigers 1–9 simple bilimbate winged capillaries.
Parapodia of abdominal chaetigers ( Fig. 7N View Figure 7 ) with spatulate to broad triangular lateral lamellae of similar size in both rami, showing a slight basal constriction, but with no obvious postchaetal expansion of lamellae behind chaetal rows. Chaetae emerging from definite ridge. Small triangular dorsal (DML) and ventral (VML) processes observed at inner margins of chaetal rows. Abdominal chaetae tridentate hooded hooks ( Fig. 7O View Figure 7 ) all of a similar size, superior two fangs parallel, above main fang. Hooks in two approximately equal groups in each ramus, main fangs vis-à-vis ( Fig. 7N View Figure 7 ). Approximately eight to ten hooks per ramus in the anterior abdomen. No abdominal pouches observed, although no posterior fragments present within examined material. Pygidium unknown.
Colour: No living material observed. Preserved specimens cream in colour, faint reddish pigment present in the posterior thorax between chaetigers 5–8 ( Figs 7A View Figure 7 , 8A, B View Figure 8 ), particularly noticeable ventrally. Pigmentation of freshly preserved specimens unknown, pigment likely to have faded to some degree after more than 10 years of preservation. Yellow dorsal, linear speckled patches (glandular?) present between chaetigers 1–4, adjacent to parapodia, either side of the mid dorsal line. Staining with methyl green ( Fig. 8C–E View Figure 8 ) shows no distinct pattern, just weak stain all over. However, dorsal speckles and abdominal interparapodial patches more apparent in stained specimens (e.g. Fig. 8C View Figure 8 ).
Distribution: Collected at 11 stations from eight countries during West African surveys at depths between 8 and 92 m. Distributed around the Gulf of Guinea, from Sierra Leone to Angola ( Fig. 1 View Figure 1 ).
Remarks: Magelona guineensis can be distinguished morphologically from all other magelonids in the MIWA region carrying posterior thoracic pigmentation using the following criteria. It differs from M. fasciata and M. alleni in possessing thoracic superior dorsal lobes, further differing from M. fasciata and, additionally, M. mackiei in possessing tridentate, instead of bidentate, abdominal hooded hooks. Magelona guineensis differs from M. alleni , M. picta and M. nanseni in the nature of the neuropodia of chaetiger 9; the neuropodial lamellae of M. alleni being ventral, as opposed to postchaetal, and in possessing additional small, triangular processes, which are not present in M. picta and M. nanseni .
The new species can also be distinguished from all other known pigmented magelonid species based on the following criteria. In stark contrast to M. guineensis , the outer pair of prostomial dorsal ridges of M. cincta are indistinct and no additional prostomial markings are present. Additionally, M. cincta does not have processes below the neurochaetae of chaetiger 9 (present in M. guineensis ), and the ventral neuropodial lamellae of the anterior thorax in M. cincta are distinctly scoopshaped (not slender triangular as in the new species). Magelona guineensis can be distinguished from M. cincta , M. equilamellae , M. japonica , M. symmetrica and M. variolamellata by possessing thoracic superior dorsal lobes. Magelona polydentata differs in possessing polydentate abdominal hooded hooks (not tridentate). Lastly, M. symmetrica is recorded to have only light pigmentation in the posterior thorax, which does not form a distinct pigment band, as seen in M. guineensis .
The only remaining African species sharing affinities with M. guineensis is Magelona cepiceps Mortimer & Mackie, 2006 described from the Seychelles. However, M. guineensis differs in the prostomial shape (being somewhat onion-shaped in M. cepiceps ), in possessing thoracic notopodial lamellae and superior dorsal lobes, which are both slender (much broader in M. cepiceps ), and in the absence of superior dorsal lobes of chaetiger 9 (present in M. cepiceps ). Magelona hartmanae Jones, 1978 , originally described off California, shares some affinities with M. guineensis in terms of lamellar and prostomial shape. However, the species differ in the nature of the postchaetal neuropodial lamellae of chaetiger 9, which is low and broad in the former species, whilst in the latter it is elongate. Additionally, the length of the prechaetal processes of the same chaetiger is much greater in M. hartmanae than the new species.
Jones ML. 1978. Three new species of Magelona (Annelida, Polychaeta) and a redescription of Magelona pitelkai Hartman. Proceedings of the Biological Society of Washington 91: 336 - 363.
Mortimer K, Mackie ASY. 2006. The Magelonidae (Annelida: Polychaeta) from the Seychelles. 2. Description of four additional species, three new to science. In: Sarda R, San Martin G, Lopez E, Martin D, George D, eds. Scientific advances in polychaete research. Scientia Marina 70: 125 - 137.

Figure 1. Map showing the MIWA sampling area between Morocco in the north and Angola in the south, indicating the position of all sampling stations and current known distributions and depths for all magelonid species carrying posterior thoracic pigmentation off the western and southern coasts of Africa.
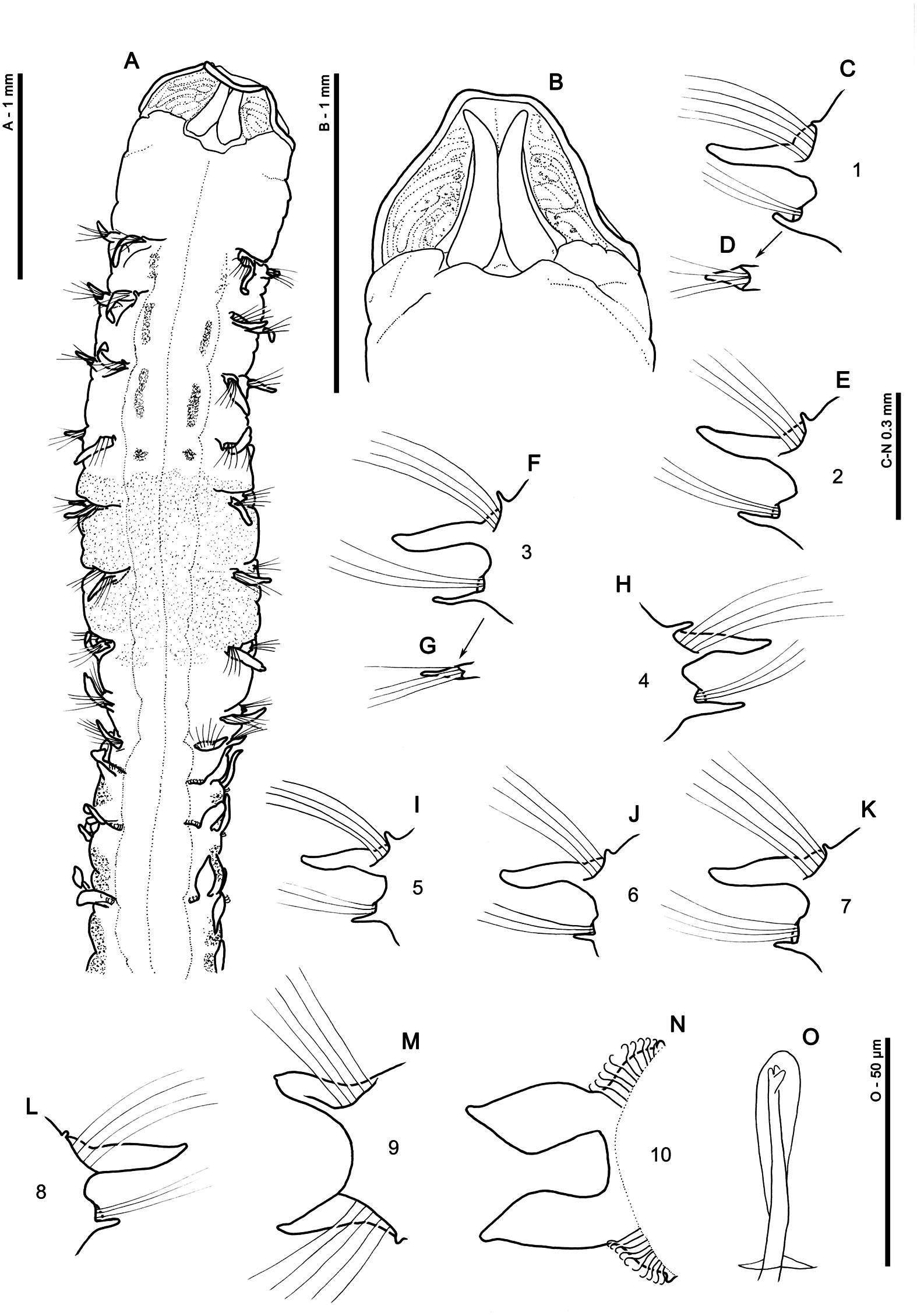
Figure 7. Magelona guineensis. Holotype (São Tomé and Príncipe, St. 5SP–05, ZMBN132137): A, anterior region (dorsal view); B, prostomium (dorsal view); C, parapodium of chaetiger 1 (anterior view); D, ventral neuropodial lamella of chaetiger 1 (dorsal view); E, parapodium of chaetiger 2 (anterior view); F, parapodium of chaetiger 3 (anterior view); G, ventral neuropodial lamella of chaetiger 3 (dorsal view); H–N, parapodia of chaetigers 4–10, respectively (anterior views); O, abdominal tridentate hooded hook (oblique frontal view).

Figure 8. Magelona guineensis. Holotype (São Tomé and Príncipe, St. 5SP–05, ZMBN132137): A, B, anterior region (dorsal and ventral views respectively, showing pigment band); C, D, anterior region (dorsal and ventral views respectively); E, prostomium (ventral view, showing buccal region). C–E, stained with methyl green.
| NMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |