
Synbranchus lampreia, Favorito & Zanata & Assumpção, 2005
|
publication ID |
https://doi.org/ 10.1590/S1679-62252005000300001 |
|
DOI |
https://doi.org/10.5281/zenodo.5070297 |
|
persistent identifier |
https://treatment.plazi.org/id/03800E70-FF9A-FF9C-FEA3-FA5807A4F936 |
|
treatment provided by |
Carolina |
|
scientific name |
Synbranchus lampreia |
| status |
sp. nov. |
Synbranchus lampreia View in CoL , new species
( Figs. 1 View Fig , 2a View Fig )
Synbranchus View in CoL sp.- Britz et al., 2003: 337-340 (larval morphology).
Holotype. MZUSP 50184 View Materials (986.0 mm TL), Brazil, Pará, ilha de Marajó, rio Goiapi, tributary of rio Arari , Cachoeira do Arari (approx. 0°58’S 48°57’W), Nov 1995, S. E. Favorito et al. GoogleMaps
Paratypes. All from Brazil, Pará, ilha de Marajó, rio Goiapi. MZUSP 50185 View Materials (4, 850.0-1.167,0 mm SL), same data as holotype GoogleMaps . MPEG 3374 View Materials (1, 857.0 mm TL), Cachoeira doArari , 20 Nov 1991, M. I. Assunção. MPEG 3377 View Materials (1, 704.0 mm TL), Taperebá, Santa Maria Farm , 23 Nov 1994, M. I.Assunção. MPEG 3379 View Materials (1, approx. 727.0 mm SL [tail damaged]), Cachoeira doArari View Materials , 23 Nov 1994, M. I. Assunção. MZUSP 84453 View Materials (3 eggs, 3.9 - 4.2 mm and 46 larvae, 13.2 –38.7 mm TL), same data as holotype GoogleMaps .
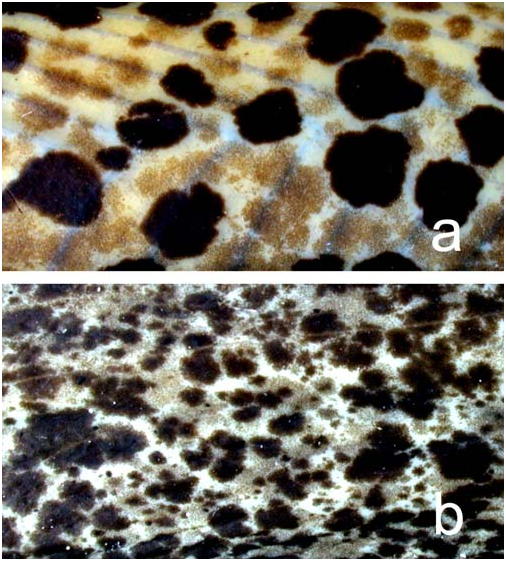
Diagnosis. Synbranchus lampreia can be easily distinguished from S. marmoratus and S. madeirae by its color pattern which consists of conspicuous large black or dark brown, roundish blotches scattered along body, rare on head, and a relatively lighter ground coloration with small and inconspicuous light brown spots ( Fig. 2a View Fig ) (vs. more homogeneously distributed spots and marbled pattern in S. marmoratus ( Fig. 2b View Fig ) and gray uniform coloration with lighter roundish spots in S. madeirae ). The new species can be further distinguished from S. marmoratus by its higher number of vertebrae (161-174 vs. 116-150) and higher number of caudal vertebrae (68-76 vs. 40- 63). Furthermore, the upper lip and the nuchal hump in S. lampreia is apparently less developed than in S. marmoratus . Synbranchus lampreia can be further distinguished from S. madeirae by its shorter post-anal length (23.2-28.7% vs. 34.7- 37.3%).
Description. Total length of examined adult specimens 704.0 to 1167.0 mm. Like that of other synbranchids, the body is slender and nearly straight on its dorsal and ventral margins for most of its length ( Fig. 1 View Fig ). Greatest body depth representing 5.3-5.7 % of total length. Body relatively long, with 161 to 174 vertebrae, of which 68 to 76 are caudal, in four specimens counted. Post-anal region compressed and relatively short. Pectoral and pelvic fins absent, the former present in larvae. Anal and dorsal-fin folds vestigial; dorsal-fin fold absent on anterior third of body, extremely narrow through most of its extension and more developed on portion situated posterior to the level of anus; anal-fin fold restricted to ventral midline posterior to anus. Caudal fin absent, without rays or hypurals. Scales absent.
Head conical, rounded anteriorly. Anterodorsal profile of head slightly concave to nearly straight; relatively pointed in lateral view. Snout short. Nuchal hump moderately developed. Eyes present, well developed, visible through a translucent layer of skin, and vertically aligned to posterior nostrils. Anterior nostrils on snout tip, anteriorly positioned; posterior nostrils situated medial to eyes. Mouth opening straight. Upper and lower jaws equal in forward extension. Premaxilla with a single series of small conical teeth. Two series of teeth on dentary and ectopterygoid similar in shape but somewhat larger and stronger than premaxillary teeth. Skin fold present over premaxilla and maxilla, forming a distinct upper lip.
Lateral line absent. Three pores of laterosensory system on dentary, two on preopercle, none on posttemporal or on parietal bones.
Branchial opening small, occupying less than one third of head width, restricted to central area of ventral surface of body, and nearly straight to crescent-shaped. Six completely ossified branchiostegal rays extending posteriorly and reaching vertical through cleithrum. Branchiostegal membrane forming folds between branchiostegal rays. Holobranchs well developed in four arches. First epibranchial without fossa to articulate with interarcual bone. Second and third basibranchials well developed. Third hypobranchial with long anteroventral process. First ceratobranchial close to first hypobranchial. Second ceratobranchial long, not expanded dorsally. Aerial suprapharyngeal sacs absent. Urohyal narrow and elongate, stalk-shaped. Posttemporal reduced to a lamellar and short stalk. Cleithrum not articulated with posttemporal, its dorsal tip extending upward at level of sixth abdominal vertebra, in lateral view. Afferent and efferent arteries associated with four branchial arches isolated and distinct from each other, but connected to dorsal aorta. Origin of internal carotid artery close to efferent artery of first branchial arch.
Color in alcohol. Ground color dark brown in dorsalmost portion of body and light brown laterally, becoming yellowish ventrally. Large black or dark brown, well-defined, roundish blotches scattered along body, more concentrated on ventral surface and on region posterior to anal opening. Large blotches usually restricted to region of body posterior to vertical through branchial opening (some blotches measuring around 18.0 mm vertically and 23.0 mm horizontally in holotype). Blotches becoming smaller on posterior portion of body. Small and inconspicuous light brown spots distributed among large dark blotches, forming a two colored blotched pattern ( Fig. 2a View Fig ). Light brown spots more conspicuous in smaller specimens and on ventral portion of body of larger individuals. These spots usually contacting each other in larger individuals, leaving just small portions of yellowish ventral ground coloration visible. Overall coloration of head similar to ground color of dorsal portion of body, without large dark blotches. Smaller specimens apparently lighter overall.
Etymology. Lampréia is a portuguese word for lamprey, used by locals at ilha de Marajó to refer to this fish. Treated as noun in apposition.
Habitat and reproductive notes. Synbranchus lampreia inhabits rio Goiapi, a white water river located on the southeastern portion of ilha de Marajó, which is situated close to mouth of the Amazon river in the northeastern Pará State ( Fig. 3 View Fig ). Ilha de Marajó, with area of 59.204 km 2, is considered one of the largest fluvio-marine islands. The type locality lies within “Campos Naturais” morphoclimatic domain, covered mainly of grass vegetation, shrubs and sparse trees (Radam, 1974). This region is influenced by periodical inundations during the rainy season, that occurs between December and June, when about 70% of the area is flooded. Rio Goiapi itself floods large areas during this period but is drastically reduced ( Fig. 4a View Fig ), and sometimes restricted to small pools during severe droughts. Nests of S. lampreia were found in dry areas situated approximately two meters away from river margin and two meters above water level ( Fig. 4a View Fig ). The nests apparently are part of a gallery system which opens externally through an usually circular opening, 10 to 15 cm in diameter ( Fig. 4b View Fig ). The external aperture leads into a tunnel running vertically downwards into the soil, for approximately 30 cm. At this point the tunnel takes a more or less horizontal direction and a floor is formed by somewhat less compact mud covered by a small pool about 2 or 3 cm deep, where eggs and larvae were found. Five nests contained 20 to 30 individuals each. Eggs and larvae in different stages of development were found to occur in the same nest (up to ten individuals) (e.g., stages A and F, described under ‘Description of eggs and larval stages’). Nests containing larvae in different stages of development were reported for S. marmoratus from the Paraguayan Gran Chaco by Breder & Rosen (1966:394, apud Taylor, 1913). These observations led us to suggest that S. lampreia eggs can be either released through many spawning activities (multiple spawning) or coexisting eggs and larvae were not necessarily offspring of the same mother (see Burt et al. (1988) for discussion about multiple spawning and Wu & Liu (1942:3) and Breder & Rosen (1966:395) for similar ideas in the synbranchid Monopterus albus ).
An adult male was always present close to the area where eggs and larvae were found. These males showed aggressive behavior as one approached the nest, suggesting some form of male parental care. Although the direction of the horizontal branch of the gallery was not exactly defined, communication between the gallery system and the Goiapi River can be assumed. Similar nests were described previously only for Synbranchus marmoratus by Taylor (1913), based on observations in the Paraguayan Chaco.
Distribution. Known only from rio Goiapi, a tributary of rio Arari, Cachoeira do Arari, ilha de Marajó, Pará, Brazil.
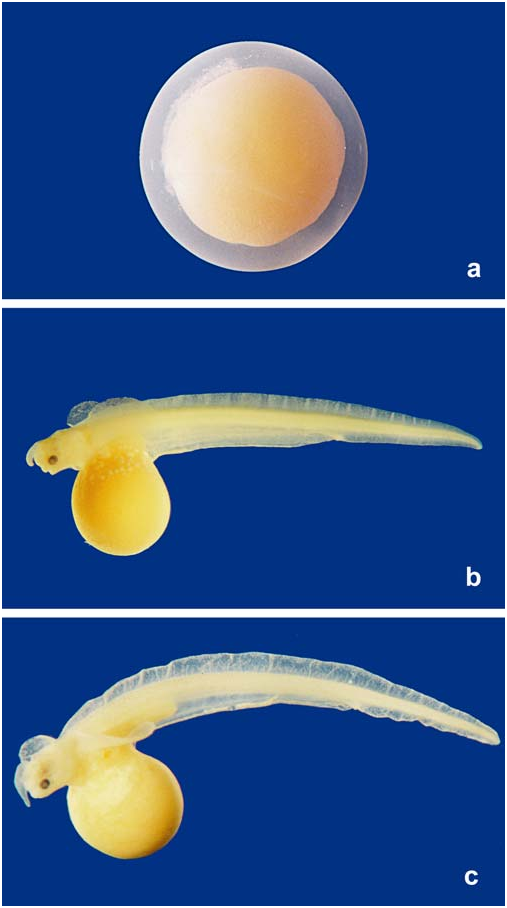
Description of eggs and larval stages. Descriptions are based on 3 eggs and 46 formalin preserved larvae measuring 13.2- 38.7 mm TL ( Figs. 5 View Fig , 6 View Fig , Table 2 View Table 2 ). Six larval stages were defined based on the degree of development of morphological characters visible externally such as pectoral fin, rostrum, and yolk inclusion. We tried to match the stages of Synbranchus lampreia with those described and illustrated by Taylor (1913) Stage B. One egg (diameter = 3.9 mm) with the embryo clearly visible through the egg capsule. At this stage there is a proboscis-like structure projecting freely from anterior end of larva (see Taylor’s fig.7, 1913). This rostrum is transparent, bluntly pointed and filled apparently with a clear fluid without any apparent cellular elements (see Figs. 5 View Fig b-c and 6a,b showing rostrum in Stages C to E). Eyes are not visible at this stage. The tail of the embryo is curled, though still in contact with the yolk. The pectorals and the unpaired larval dorsal and anal median fin folds are already visible under the capsule and are also curled. They extend as relatively translucent membranes of skin from the point of pectoral-fin insertion to the posterior tip of the curled tail. The depth of larval dorsal and anal fin folds are similar, and equivalent to body depth. Distance from anterior tip of rostrum to anterior end of encephalon 0.95 mm, from anterior tip of rostrum to pectoral-fin insertion 2.5 mm, pectoral-fin length 3.3 mm. Stage B described herein probably corresponds to Taylor´s Stage 23 (1913).
for S. marmoratus . The length of some measurements in the larvae we described are longer than those of S. marmoratus , a fact probably related to the relative larger size of the adults of S. lampreia (704.0-1167.0 mm TL versus maximum TL of 857.0 mm in S. marmoratus ).
Stage A. Two spherical eggs (diameter = 4.1 and 4.2 mm) with embryos not visible externally ( Fig. 5a View Fig ). The eggs consist of a firm and somewhat tough capsule separated from the yolk by the perivitelline space.A mass of what looks like a coagulated albuminous substance is present at the vegetative pole, similarly to that described for eggs of S. marmoratus by Taylor (1913:4). This stage probably corresponds to Taylor’s (1913:7) Stage 17 with a germinative pole containing a Y-shaped white line of 1.9 mm (compare with Taylor’s fig. 4).
Stage C. Three hatched embryos with main portion of body completely free from yolk sac and without significant yolk included in abdominal cavity ( Fig. 5b View Fig ).After intermediate stage of hatched and coiled embryo, between Stage B and C not documented herein, the coiled portion of body straightens out as the embryo grows, so that at stage C a straight line joining tip of rostrum to tip of tail measures 13.2-15.3 mm. The rostrum has lengthened and lies curled in parallel with the curve of the yolk-sac. Although without clearly defined pigmentation, a large and laterally positioned eye is visible. The mouth is well defined and the opercular opening is clearly visible. Larval anal-fin fold somewhat deeper than dorsal-fin fold, its depth measuring 0.5 mm in the area close to the yolksac. Segmentation of musculature clearly visible. Probably correspond to or is slightly more developed than Taylor´s Stage 26 (1913:11, fig. 9).
Observations of live larvae at this stage, and the next two stages, showed a dense network of capillaries in the pectoral fins. Active movements of these fins propelled water from a well-circumscribed area anterodorsal around the head to the posterior portion along the length of the entire larva and its yolk sac. These pectoral fin movements were previously described for larvae of Synbranchus marmoratus ( Taylor, 1913:15) , Monopterus cuchia (Banerj et al., 1981:149; Munshi et al., 1989a:310), and Monopterus albus (Wu & Liu, 1942:6; Liem, 1981:1177). According to Liem, synbranchid larvae can survive periods of critical hypoxia because they have a combination of adaptations, including highly vascularized pectoral fins and other large vascularized areas of body, an active pectoral fin mechanism that generates a posteriorly directed respiratory water current originating from the oxygen-rich surface layer, allied with a small blood-water barrier, and a principal flow of blood that runs in form of a countercurrent to the water stream. As described in item “Habitat and reproductive notes” eggs and larvae of S. lampreia were collected in 2 or 3 cm small and poorly oxygenated pools, located distant from stream water. Presence of pectoral fins is restricted dorsal and ventral musculature visible as a longitudinal sulcus. Dorsal larval fin fold apparently deeper in smaller specimens, becoming narrower in larger ones. Pectoral fin similar to that described in the previous stage, although comparatively larger. Dark chromatophores are present on dorsal portion of head and body, starting immediately posterior to rostrum and becoming densely concentrated in a horizontal stripe along dorsal-fin fold base. Eyes pigmented. Probably corresponds to Taylor´s Stage 30 (1913:15, fig. 10).
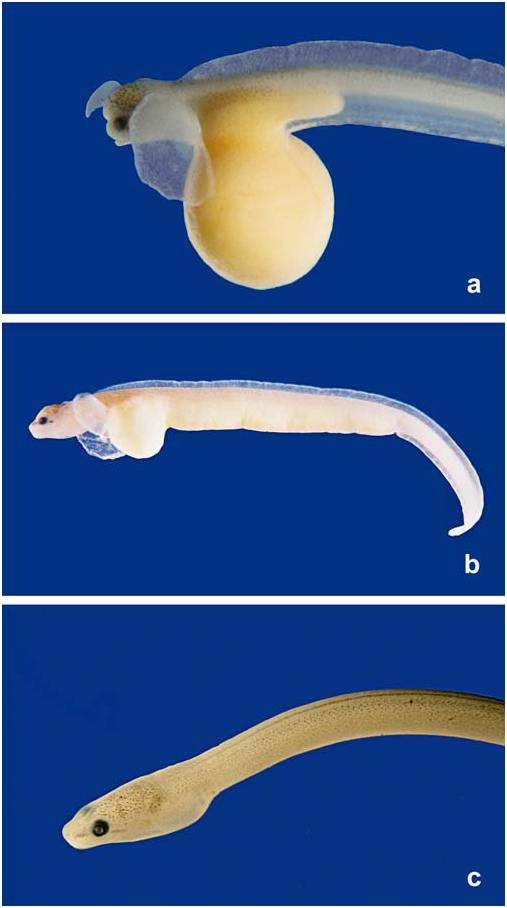
Stage E. Ten larvae (4 measured, 21.5-23.9 mm SL) with large amounts of yolk enclosed in the abdominal cavity ( Fig. 6b View Fig ). The yolk enclosed in the embryo´s body extends to within 1.0 mm of the anus in larger specimens examined and has total extension of about 11.5-15.2 mm. The yolk sac has almost lost its globular appearance. Rostrum very reduced, absent as a free structure and tied to the dorsal portion of the head. Pectoral fin reduced in proportion to rest of body and more ventrally positioned. Anal larval fin fold restricted to portion of body posterior to anal opening. Post-anal portion somewhat curled ventrally.
At this stage the larvae are more densely pigmented. Dorsal and lateral surfaces of head with inconspicuous dark chromatophores, reaching approximately the horizontal through center of eye and continuing dorsally and posteriorly through dorsal-fin fold base. Lateral and ventral portion of body and fins with sparsely distributed dark chromatophores. Presence of conspicuous and horizontally elongated dark stripe, starting anterior to eye and continuing posteriorly beyond it for an extension approximately equal to eye diameter. Probably corresponds to Taylor´s Stage 32 (1913:16, fig. 13).
to stages described herein as C, D and E, being completely absent in Stage “F” when the branchial chamber has the respiratory function.
Stage D. Eleven larvae (15.1-19.6 mm TL) with a certain degree of yolk inclusion in their abdominal cavities ( Fig. 5c View Fig and 6a View Fig ). Although assigned to just a single stage, these larvae show some variation in the amount of egg inclusion and in rostrum size and position. Largest portion of rostrum (2/3) situated over head and smaller anterior portion (1/3) is hanging free anterior to head; rostrum terminal in the smaller specimens (those with less amount of included yolk) becoming shorter and more anterodorsally positioned in larger specimens. Myomeres well defined. Demarcation line between Stage F. Twenty-two specimens with body shape similar to adults (35.0- 38.7 mm TL) ( Fig. 6c View Fig ). The specimens are eel-like shaped, the body behind opercular cavity being almost perfectly cylindrical, while the posterior part of the trunk becomes gradually laterally compressed and merges into the tail. There is no vestige of pectoral fins and no trace of rostrum. The yolk sac is completed absorbed. Lateral portion of branchial opening completely closed with single aperture limited to ventral portion of body. Chromatophores are still more conspicuous on the dorsal and lateral portions of head and body where they are situated close to the skin surface. Dark stripe anterior to eye still present. Chromatophores situated on ventral half of body inserted deeply in skin, giving a faded appearance to pigmentation. Chromatophores situated on lateral portion of body arranged along the segmented limits of the V-shaped myomeres. Ventral midline without pigmentation. Corresponds to Taylor´s Stage 35 (1913:18, fig. 16).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Synbranchus lampreia
| Favorito, Sandra E., Zanata, Angela M. & Assumpção, Maria I. 2005 |
Synbranchus
| Britz, R 2003: 337 |