
Spirobranchus tetraceros ( Schmarda, 1861 )
|
publication ID |
https://doi.org/10.11606/1807-0205/2020.60.51 |
|
publication LSID |
lsid:zoobank.org:pub:C598BF30-2A01-467A-B4F1-42032F448100 |
|
DOI |
https://doi.org/10.5281/zenodo.4975233 |
|
persistent identifier |
https://treatment.plazi.org/id/025CD000-8802-9C63-FC39-47AAFC41F8BE |
|
treatment provided by |
Carolina |
|
scientific name |
Spirobranchus tetraceros ( Schmarda, 1861 ) |
| status |
|
Spirobranchus tetraceros ( Schmarda, 1861) View in CoL
( Figs. 7 View Figure 7 , 8 View Figure 8 , 9 View Figure 9 )
Pomatoceros tetraceros Schmarda, 1861 .
Spirobranchus tricornis View in CoL . Straughan (1967).
Spirobranchus tetraceros View in CoL . ten Hove (1970).
Spirobranchus dendropoma View in CoL . ten Hove (1970).
Spirobranchus giganteus View in CoL . Dew(1959); Skinner et al. (2012).
Complex- Spirobranchus tetraceros View in CoL . ten Hove (1994).
Examined material: 28 specimens. Rio de Janeiro state, Ilha Grande Bay, Angra dos Reis Municipality: Cataguases Island: 23°01′28.57″S, 44°17′06.52″W, intertidal, MNRJP-002787 ( 2 specimens). Paraty Municipality: Catimbau Island: 23°11′42.41″S, 44°37′33.19″W, UERJ 6284 ( 1 specimen). Mangaratiba Municipality: Ibicuí Beach: 22°57′45.30″S, 44°01′28.50″W, MNRJP-002788 ( 10 specimens). Itaguaí Municipality: Coroa Grande Beach: 22°54′24.34″S, 43°52′00.26″W, MNRJP-002789 ( 15 specimens).
Diagnosis (after Kupriyanova et al., 2015): Opercular plate mostly flat, sometimes conical; basally carrying 1 mid-ventral and 2 dorso-lateral spines (“tricornis” condition); alternatively, some specimens with mid-ventral spine as two fully separated spines, which, considering pair latero-dorsal spines, makes 4 primary spines (“tetraceros” condition). All spines split at least once, frequently twice or three times, resulting in 6-14 spine tips. Completely conical opercula, with only 3 rudimentary spines or spines completely absent. Peduncular wings with finger-like processes distally (rallely without).
Description
Tube: Opaque, white or pinkish. Triangular in cross-section, lumen circular. One serrated longitudinal keel throughout, extending over the opening as pointed tip ( Fig. 7A View Figure 7 ). Tubes solitary.
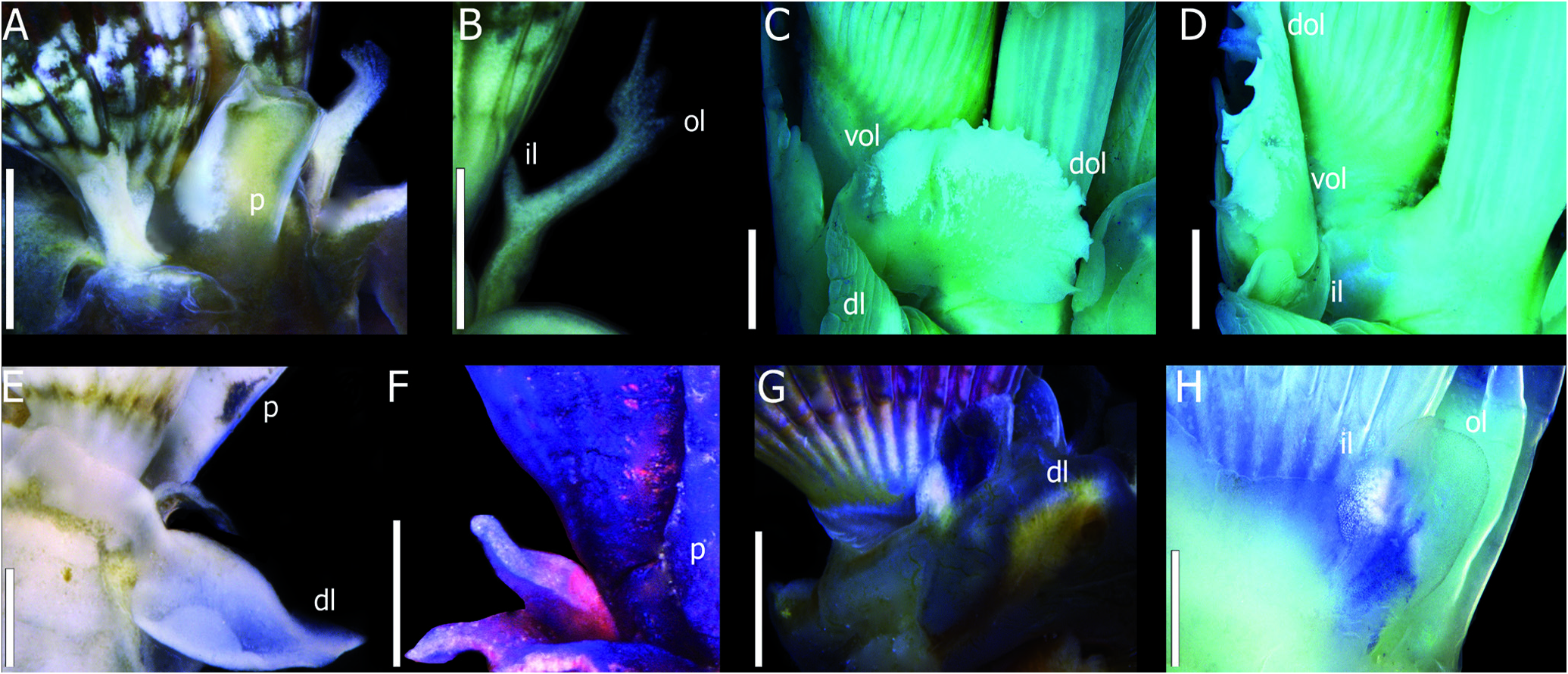
Radiolar Crown: Live specimens alternating bands of white iridescent, yellow, brown, pink, and orange coloration at crown ( Fig. 7B View Figure 7 ). Coloration faded after preservation, only darker shades remaining. Crown as two spiraled lobes with 1 whorl, each with up to 27 radioles, basally joined by inter-radiolar membrane for one third of their length; inter-radiolar membrane bearing triangular distal processes, with fringed edges ( Fig. 7C View Figure 7 ), usually white iridescent ( Fig. 7H View Figure 7 ). Radioles rectangular in cross-section, inner side with two rows of filiform pinnules of even length through radioles, with naked tips. Naked tips filiform and about as long as pinnules. Dorsal radioles longer than ventral ones.Scattered simple ocelli, in irregular rows through radioles.
Mouth Parts: Two thin, smooth lips. Dorsal lips rectangular, forming pair of lateral cylindrical grooved palps, basally flat, about half as long as dorsal radioles ( Fig. 7I View Figure 7 ). Ventral lip rectangular, about as long as dorsal lips, as internal lap through bases of radiolar lobes.
Peduncle: Smooth, triangular in cross-section, inserted slightly left to dorsal mid-line. Pair of smooth lateral wings distally, bilobed, dorsal lobe circular, lateral lobe triangular, both with fringed edges ( Fig. 7F View Figure 7 ). Constriction between peduncle and opercular base absent. Darkbrown, alternating white iridescent bands often present, with scattered pigmentation, as in crown. Coloration pattern usually extending into lateral wings.
Operculum: Continuous with the peduncle. Opercular ampulla flat, circular to oval in top view, calcareous endplate usually white or beige. Endplate varying from a ridged conical ( Fig. 7D View Figure 7 ) structure to a flat plate with three main spines ( Fig. 7E View Figure 7 ), two dorso-lateral and one medio-ventral, wich may split at base, resulting in four main concentric spines ( Figs. 7F,7G View Figure 7 ). Spines covered by spaced smaller spines. Multiple branching in each spine resulting in many degrees of development of spines observed in a population.Talon absent.
Collar and Thoracic Membranes: Three-lobed collar. Dorso-lateral lobes longer, circular, with entire edges, continuous with thoracic membranes, forming a ventral apron covering 1 abdominal chaetiger ( Fig. 7H View Figure 7 ). Midventral lobe rectangular with pointed median tip, shorter than dorso-lateral lobes, often folded medially, inserted between crown lobes.Tonguelets present between lateral and ventral lobes; bilobed, inner lobes half-length of outer lobes, both circular, with smooth rim ( Figs. 7H View Figure 7 , 9G, 9H View Figure 9 ). Collar lobes commonly same color as crown base, extending through thorax. Internally, collar and tonguelets may be covered in white iridescent pigmentation.
Thorax: Seven chaetigers. Collar chaetae as Spirobranchus - type and limbate chaetae, disposed in one fascicle ( Fig. 8L View Figure 8 ). After collar, notopodial lobes round, emerging between thoracic membranes and neuropodia. Thoracic chetae limbate, in three fascicles of different lengths per notopodium ( Figs. 8M, 8N View Figure 8 ). Neuropodial lobes rectangular, all bearing tori curved downwards of even length through thorax.Thoracic uncini saw-shaped, with 13-16 curved teeth, and gouged anterior peg ( Fig. 8O View Figure 8 ). Parapodial lobes approaching each other posteriorly, forming ventral triangular depression. Glandular clusters covering ventral portion of mid-ventral collar lobe, as heart-shaped pattern, together with triangular depression ( Figs. 7J, 7K View Figure 7 ), also covering all thoracic neuropodia, except for origin of tori. Five pairs of ventral glandular shields, first two pairs rectangular, fused to each other mid-ventrally, other three pairs not fused and posteriorly becoming squared ( Figs. 7J, 7K View Figure 7 ).
Abdomen: Largest analyzed specimen with 97 abdominal segments. Abdomen in entire animals usually four times longer than thorax. Notopodial uncini smaller than thoracic ones, saw-shaped,with 10-11 teeth and gouged anterior peg ( Fig. 8P View Figure 8 ). Neuropodial chaetae true trumpet-shaped, abruptly bent, with 2 distal rows of denticles separated by groove and lateral serrated filiform projection ( Fig. 8Q View Figure 8 ). Ventral pads square-shaped anteriorly and rectangular posteriorly. Glandular clusters present over tori, and along the rim of fecal sulcus, concentrated at posterior abdomen. Pygidium bilobed, with terminal anus.
Remarks: The occurrence of S. tetraceros in the Atlantic has been deemed indicative of a species complex ( Smith, 1985; Fiege & ten Hove, 1999; ten Hove & Kupriyanova, 2009). This species was first described in Australia ( Schmarda, 1861) and its members are similar to those of the Caribbean S.dendropoma Mörch,1863 . Spirobranchus dendropoma has been treated as a synonym of S. tetraceros ( e.g., ten Hove, 1970), but that status is questionable, given the improbability of an Indo-Pacific species occurring in the Caribbean ( Bastida-Zavala & Salazar-Vallejo, 2000; Bastida-Zavala, 2009; Perry et al., 2018). However, morphologically, members of both these species differ in terms of the processes present at the distal edge of the interradiolar membrane, which are smooth among specimens of S. dendropoma , and positioned at the base of the radioles ( Benedict, 1887), whereas they are fringed in animals belonging to S. tetraceros , and at the distal end of the interradiolar membrane (Bastida-Zavala & Salazar-Vallejo, 2000). Perry et al. (2018) alert that S. tetraceros is an extensively distributed complex of species, though that do not exclude the likelihood of either of these species being introduced distant to the type-locality. The specimens we analyzed are compatible with descriptions of S. tetraceros , and the occurrence of this exotic species in Brazil was recently noticed ( Rodrigues et al., 2020), thus we retain the identification of the mentioned previous authors.
Members of S. tetraceros share with those of S. giganteus the tube with a pointed longitudinal keel, operculum ornamented by thorns, and special collar chaetae. However, the tube of members of the former species may present serrations on the keel, which are absent in S. giganteus . The radiolar lobes of individuals of S. giganteus form multi-whorled spirals, whereas they form a single circle in members of S. tetraceros . The opercular thorns of specimens of S. tetraceros are more numerous, up to four, concentric and branched, with multiple tips, while in animals of S.giganteus there are two dorso-lateral and a possible third mid-ventral thorn, shorter, in a triangular arrangement with other thorns on the opercular endplate. The tonguelets of members of S. tetraceros are bilobed with smooth edges, whereas specimens of S. giganteus present trilobed tonguelets, with superior outer lobe larger and with wavy edge. The peduncular wings and interradiolar processes of animals of S. tetraceros are fringed, while in members of S. giganteus the wings are smooth, and the processes are absent. Members of S. tetraceros have 5 pairs of glandular shields in thoracic triangular depression, while those of S. giganteus have a single shield.
Spirobranchus tetraceros differs from S. minutus because members of S. minutus have a triple-keeled tube, with alveoli,lack both opercular ornamentation and radiolar ocelli, and the collar chaetae, if present, are limbate, while in members of S. tetraceros the tube has a single keel, the operculum is ornamented,photoreceptor structures are present and the collar chaetae are limbate and special Spirobranchus - type.
Members of S.tetraceros share with S.lirianeae sp.nov. the shape of the tube, with single pointed longitudinal keel, but they differ from those animals in having opercular ornamentation, simple radiolar ocelli, fringed interradiolar processes, and collar chaetae, while members of S. lirianeae sp. nov. have funnel-shaped operculum, lacking thorns and spines, and also have paired compound ocellar clusters in the radioles and interradiolar processes, whereas collar chaetae are absent. The fringed peduncle wings and the presence of interradiolar processes distinguish this species from any other recorded in Brazil to date.
Type-locality: New South Wales, Australia (Pacific Ocean).
Habitat: Intertidal: rocky shores, beneath rocks; subtidal: rocky shores, under rocks, at fouling communities on piers. Often found associated to Tubastraea spp. (Cnidaria: Anthozoa: Scleractinia ). Also found on artificial substrata, such as concrete blocks and plastic incrustation plates ( Rodrigues et al., 2020).
Distribution: Indian Ocean: Australia, Phillipines, off China ( Sun et al., 2012), Red Sea ( Perry et al., 2017),Aqaba Gulf,eastern coast of Africa and Madagascar ( Read,2019); Pacific Ocean: Australia; Atlantic Ocean: Mediterranean Sea, Greek, Caribbean Sea ( ten Hove, 1970), Gulf of México, Colombia ( Read, 2019) and southeastern Brazil, at Rio de Janeiro state ( Skinner et al., 2012; Rodrigues et al., 2020).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Spirobranchus tetraceros ( Schmarda, 1861 )
| Brandão, Ivison Cordeiro & Brasil, Ana Claudia dos Santos 2020 |
Spirobranchus tricornis
| Morch 1863 |
Spirobranchus dendropoma
| Morch 1863 |
Pomatoceros tetraceros
| Schmarda 1861 |