
Bryodelphax instabilis, Gąsiorek & Degma, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4410.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:0944C1F0-1405-43E0-80B7-03438A19F334 |
|
DOI |
https://doi.org/10.5281/zenodo.5998748 |
|
persistent identifier |
https://treatment.plazi.org/id/00108791-0D06-FF9A-D399-FE75FB44F80C |
|
treatment provided by |
Plazi |
|
scientific name |
Bryodelphax instabilis |
| status |
sp. nov. |
Bryodelphax instabilis View in CoL sp. nov.
( Figs 1–23 View FIGURES 1–5 View FIGURES 6–9 View FIGURES10–12 View FIGURES13–16 View FIGURES 17–18 View FIGURES 19–20 View FIGURES21–23 , Tables 3–5)
Locus typicus. Moss on carbonate bedrock from Homole Ravine in the Pieniny Mts. ( 49°24'17''N, 20°32'52''E; 595 m asl), Poland. GoogleMaps
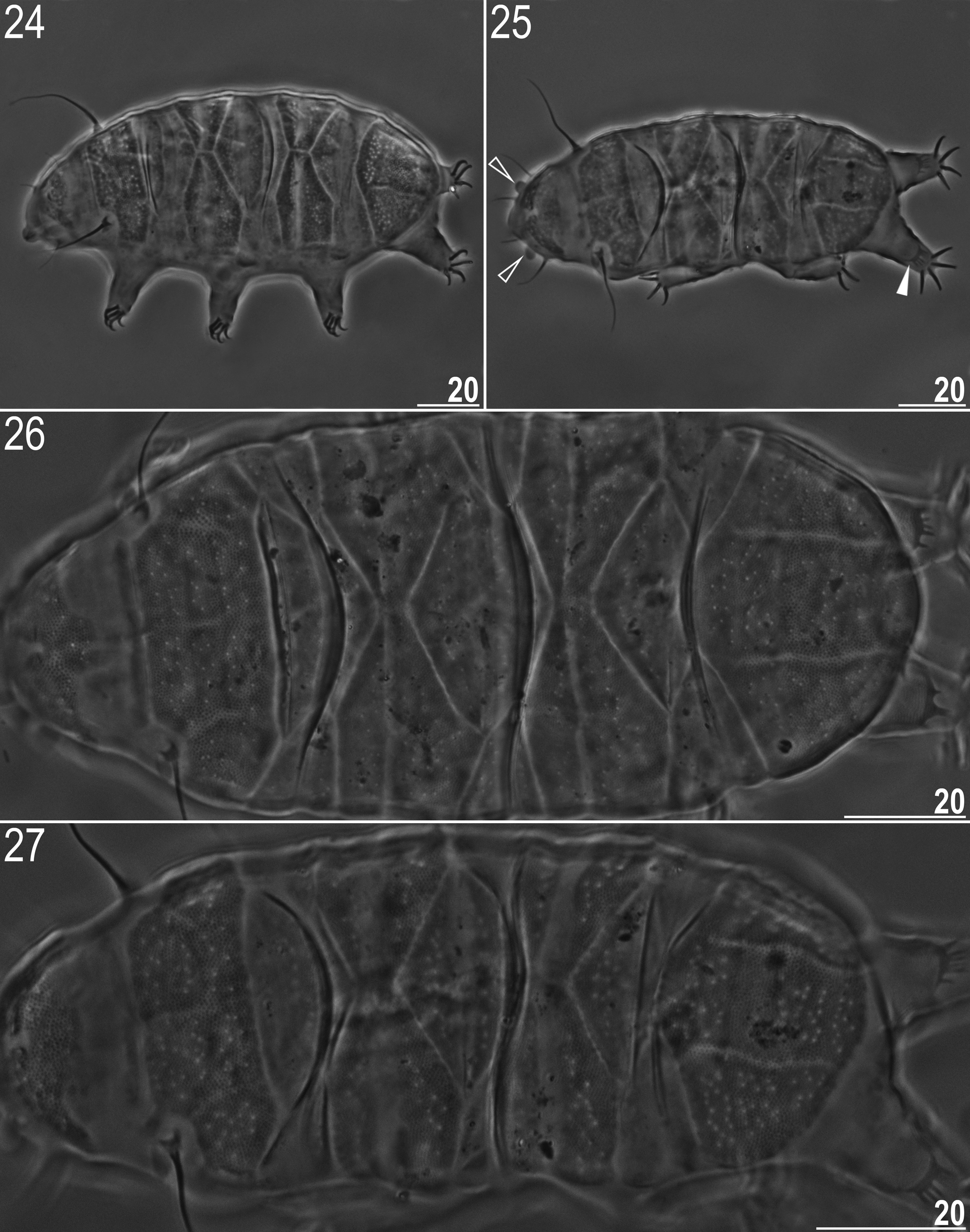
Type material. Holotype (mature female, slide no. PL.189.19), coll. Maciej Barczyk, 25th May 2016, allotype (mature male, slide no. PL.189.01), 30 paratypes (21 mature females, 7 males, one juvenile, and one larva; slides PL.189.02–32) and additional 15 paratypes (both juveniles and adults) on SEM stubs. Found together with numerous Bryodelphax parvulus Thulin, 1928 ( Fig. 24 View FIGURES 24–27 ), Pseudechiniscus suillus ( Ehrenberg, 1853) , and single specimens of Testechiniscus spitsbergensis ( Scourfield, 1897) . All from the same moss sample. Holotype and most of paratypes deposited in the Institute of Zoology and Biomedical Research, Jagiellonian University, Kraków, Poland, two paratypes (slides PL.189.10–11) deposited in the Department of Zoology, Comenius University in Bratislava, Slovakia, and two paratypes (slides PL.189.22–23) deposited in the Zoological Museum, University of Copenhagen, Denmark.
Additional material examined. Two specimens in a moss sample from Chuda Alp in the Tatra Mts. (slide PL.258.06), coll. Piotr Gąsiorek, 24th September 2016, deposited together with type material. 128 specimens ( 44 males, 43 females, 3 juveniles, and 38 specimens of unidentified sex due to unsuitable orientation of a specimen on a slide; slides 390/7–10, 390/18, 390/20, 390/23, 390/25, 390/27–30, and 390/32; deposited in the Department of Zoology , Comenius University in Bratislava , except for 13 specimens deposited in the Institute of Zoology and Biomedical Research , Jagiellonian University , Kraków — slide 390/18) and 9 specimens used for SEM analyses (not deposited) in a moss sample on a rock from Rajtopíky hill, the Branisko Mts. ( 48°59'42''N, 20°51'59''E; ca. 1036 m asl — on the hill top area), Eastern Slovakia, coll. Peter Degma, 26th July 2003 GoogleMaps .
Short description of holotype ( Figs 1 View FIGURES 1–5 , 10 View FIGURES10–12 ; Table 3)
Adult female; colour greyish; eyes not visible (after preparation); body length 137 µm ( 685); cephalic cirri and papillae, and clava not enlarged; cirrus A 26.5 µm ( 132.5); claws I–III of similar size, claw IV longer; dentate collar present, with three teeth; small papilla on leg IV. Dorsal plates with sparse pseudopores/pores; ventral plates not obvious. Female gonopore rosette-shaped.
Short description of allotype ( Figs 2 View FIGURES 1–5 , 12 View FIGURES10–12 ; Table 4)
Adult male; colour greyish; eyes not visible (after preparation); body length 123 µm ( 628); cephalic cirri and papillae, and clava enlarged; cirrus A 20.7 µm (103.0); claws I–III of similar size, claw IV longer; dentate collar present, with three teeth; small papilla on leg IV not visible. Dorsal plates with sparse pseudopores/pores; suggestion of deeper faceting on scapular and caudal plates; ventral plates present. Male gonopore simple, circular.
Details of the new species. Adults ( i.e. from the third instar onwards; measurements and statistics for ♀♀ in Table 3, for ♂♂ in Table 4):
Body greyish; eyes absent or not visible after preparation. Mouth opening surrounded by a ring of 10 papulae (visible in SEM only). Cephalic papillae greatly enlarged in males (compare Figs 1 and 2, 4 View FIGURES 1–5 ), and clavae are also more prominent ( Figs 13 View FIGURES13–16 , 17–18 View FIGURES 17–18 ). Internal cirri much shorter than external ( Tables 3–4, Figs 1–4 View FIGURES 1–5 , 7–9 View FIGURES 6–9 ). Cirri A thinner at the tip (visible only under SEM; Fig. 17 View FIGURES 17–18 ). Dorsal plates covered with sparsely distributed pseudopores or pores of large and intermediate size (visible in PCM as bright dots with margins either blurred (pseudopores) or obvious (pores); Figs 1–2 View FIGURES 1–5 , 10, 12 View FIGURES10–12 ), true pores occur infrequently, being restricted to the posterior portion of the cephalic plate and the anterior portion of the scapular plate ( Fig. 13 View FIGURES13–16 ) in the proximal body part, to the posterior parts of segmental plates ( Fig. 14 View FIGURES13–16 ), and the anterior margin of the caudal plate in the distal body part. Pores rarely present on all plates ( Figs 11 View FIGURES10–12 , 15–16 View FIGURES13–16 ), but are often absent ( Figs 6–7 View FIGURES 6–9 ). Pseudopores/pores density similar on all dorsal plates, approximate mean density 10–13 pseudopores/pores per 100 µm 2 in females and 8–10 pseudopores/ pores per 100 µm 2 in males ( Table 5). Intracuticular pillars visible as fine dark dots under PCM ( Figs 10–12 View FIGURES10–12 ). Scapular plate typically distinctly faceted with a median longitudinal fold and three smaller transverse folds ( Figs 2 View FIGURES 1–5 , 12 View FIGURES10–12 ). In females, sometimes only pseudopore/pore rows mark borders of faint facets ( Figs 1 View FIGURES 1–5 , 10–11 View FIGURES10–12 ). Paired plates divided into two unequal anterior and posterior parts by a transverse stripe ( Figs 10–12 View FIGURES10–12 , 14, 16 View FIGURES13–16 ). Caudal plate faceted with two evident longitudinal folds ( Figs 1–2 View FIGURES 1–5 , 6 View FIGURES 6–9 , 10–12 View FIGURES10–12 ), most often the central portion of this plate is subdivided into two parts by a transverse V-suture ( Figs 2 View FIGURES 1–5 , 6 View FIGURES 6–9 , 12 View FIGURES10–12 ). Median plates 1 and 2 with transverse division into two unequal parts (compare Figs 10–12 View FIGURES10–12 , 14, 16 View FIGURES13–16 ). Median plate 3 with faint transverse suture, triangular in shape, and with a roundish posterior edge ( Figs 14, 16 View FIGURES13–16 ). Poorly developed supplementary lateral platelets present at the levels of median plates (three pairs of platelets on each body side: a pair between scapular plate and first pair of the segmental plates, a pair between paired plates, and a pair between second pair of segmental plates and caudal plate; Figs 1 View FIGURES 1–5 , 10–12 View FIGURES10–12 ), devoid of pores/pseudopores ( Fig. 7 View FIGURES 6–9 ). Ventral cuticle typically with weakly developed plates. Ventral intracuticular pillars either absent or present and well-visible (compare Figs 4–5 View FIGURES 1–5 ) at 100x oil immersion, and always less evident than dorsal (compare Figs 5 View FIGURES 1–5 and 11 View FIGURES10–12 ). Ventral plate configuration: VII/IX:(2)-(1)-2/4-2-2/4- 2-2-2-1. Two small subcephalic longitudinal plates just below the mouth opening and one subpharyngeal plate weakly outlined and usually only visible under SEM (compare Figs 5 View FIGURES 1–5 , 8–9 View FIGURES 6–9 ). Typically, rows III–VIII/IX are discernible ( Figs 4–5 View FIGURES 1–5 , 8–9 View FIGURES 6–9 , 21 View FIGURES21–23 ). Rarely, ventral plates invisible under PCM ( e.g. holotype). Ventral granules absent or restricted to the plate surface ( Fig. 5 View FIGURES 1–5 ). Papilla on legs I absent, minute papilla on legs IV present ( Fig. 1 View FIGURES 1–5 ). Dentate collar on legs IV with 3–6 teeth ( Figs 1–2 View FIGURES 1–5 , 20 View FIGURES 19–20 , arrowheads). External claws of all legs smooth, internal claws with tiny spurs pointing strongly downwards with very small gap between spur and claw base, making them almost invisible under PCM (however, always clear under SEM; Figs 19–20 View FIGURES 19–20 ).
Remarks on the sexual dimorphism: Sex differences in the new species are well-marked and embrace: longer, tubbier primary and secondary ♂ clavae (compare Figs 1–2 View FIGURES 1–5 and Tables 3–4); stronger faceting of the ♂ scapular plate (compare Figs 10–11 and 12 View FIGURES10–12 ); significantly lower pseudopore/pore density on unpaired plates in ♂ (compare ranges presented in Table 5).
Juveniles ( i.e. the second instar, four-clawed without gonopore):
In appearance like adults, but clearly smaller (75 µm) and with indiscernible ventral plates ( Fig. 22 View FIGURES21–23 ). It is impossible to state firmly whether the species exhibits ontogenetic shifts in ventral armature or the lack of ventral plates in the first two life stages is inconstant. This issue certainly requires more studies, since changes in ventral armature occurring during development were recently detected by Gąsiorek et al. (2017).
Larvae ( i.e. the first instar, two-clawed without gonopore; measurements in Table 4):
Median plates with poorly delineated margins, lacking pseudopores/pores; supplementary lateral platelets and pedal (leg) plates undeveloped. Pores/pseudopores present only on the anterior and posterior margins of the cephalic, scapular, segmental, and caudal plates ( Fig. 3 View FIGURES 1–5 ). Claws with spurs formed as in adults. Legs IV without dentate collar. Ventral plates absent ( Fig. 23 View FIGURES21–23 ).
Eggs unknown.
Etymology: The name instabilis underlines the variability in the number of ventral plates (see Remarks), which is unusual for Bryodelphax . Moreover, often only some of the ventral plates rows are visible under the light microscope, thus an animal’s venter seems to be devoid of plates in its proximal part.
Remarks. Polish and Slovak populations have same ventral plates configuration but in three mature females from the Slovak population, row V has two additional, smaller ventral plates placed in a more marginal position, and two of the specimens also have such additional plates in row III ( Figs 5 View FIGURES 1–5 and 21 View FIGURES21–23 ).
Differential diagnosis. Bryodelphax instabilis sp. nov. can easily be distinguished from the other members of the weglarskae group on the basis of the scapular plate faceting and being gonochorous. Nevertheless, the new species should be compared with four most similar taxa having last six ventral plates rows configuration same as the new species, i.e. 2-2-2-2-2-1 or 2-4-2-2-2-1 (due to the instability in ventral plate arrangement). It differs specifically from:
Bryodelphax iohannis Bertolani et al., 1996 View in CoL , by a different ventral plate configuration (VII/IX:(2)-(1)-2/4-2-2/ 4-2-2-2- 1 in the new species vs X: 2-1-1-5-2-4-2-2-2- 1 in B. iohannis View in CoL ), by the presence of lateral platelets (absent in B. iohannis View in CoL ) and by longer teeth on the dentate collar (compare Figs 1–2 View FIGURES 1–5 with Fig. 2A View FIGURES 1–5 in Bertolani et al. 1996);
Bryodelphax parvuspolaris Kaczmarek et al., 2012 View in CoL , by a different ventral plate configuration (VII/IX:(2)-(1)- 2/4-2-2/4-2-2-2- 1 in the new species vs VIII: 1-1-2-2-2-2-2- 1 in B. parvuspolaris View in CoL ), and by well-developed dentate collar (dentate collar with poorly developed teeth in B. parvuspolaris View in CoL );
Bryodelphax sinensis ( Pilato, 1974) View in CoL , by a different ventral plate configuration (VII/IX:(2)-(1)-2/4-2-2/4-2-2-2- 1 in the new species vs VII:2-2-2-2-2-2- 1 in B. sinensis View in CoL ), and by the presence of well-developed dentate collar on legs IV in adults (dentate collar absent in B. sinensis View in CoL );
Bryodelphax weglarskae ( Pilato, 1972) View in CoL , by a different ventral plate configuration (VII/IX:(2)-(1)-2/4-2-2/4-2- 2-2- 1 in the new species vs IX:2-1-5-2-4-2-2-2- 1 in B. weglarskae View in CoL ), and by non-bifurcated cephalic appendages (bifurcated in B. weglarskae View in CoL ).
Moreover, Bryodelphax instabilis View in CoL sp. nov. must be primarily compared with the first described gonochoristic representative of the genus, namely Bryodelphax tatrensis (Węglarska, 1959) View in CoL because it resembles the latter species when the ventral armature is faint. Bryodelphax instabilis View in CoL sp. nov. is distinguished from B. tatrensis View in CoL on the basis of: (1) the average pseudopore/pore size is larger in the new species in comparison with the minute pores in B. tatrensis View in CoL (the difference is especially obvious between females of both species, compare Figs 10–11 View FIGURES10–12 and 26 View FIGURES 24–27 ), (2) more pronounced faceting of the scapular plate (clearly visible median suture and typically well-developed facets in the new species vs only faint median suture, which can be absent in B. tatrensis View in CoL ; compare Figs 11–12 View FIGURES10–12 and 26– 27 View FIGURES 24–27 ), and (3) the caudal plate consisting of four facets in males of the new species instead of three in B. tatrensis View in CoL males (compare Figs 2 View FIGURES 1–5 , 12 View FIGURES10–12 and 25, 27 View FIGURES 24–27 ). Additional discriminative criteria ( e.g. morphometric) could be presented when measurements of a large population of B. tatrensis View in CoL become available. Unfortunately, general rarity of B. tatrensis View in CoL hinders the redescription of this taxon ( Dastych 1988, personal observations).
General remarks on Bryodelphax in Polish Pieniny Mts. and Tatra Mts. There are four sympatric Bryodelphax spp. that inhabit Poland and occur in the Pieniny Mts. ( Dastych 1988); three (with the exception of B. weglarskae ) are also present in the Tatra Mts. Our new discovery confirms an eucalciphil and subalpine preference for these Central European members of the genus. In Poland, Bryodelphax spp. are frequently found in close proximity. For example, in the Pieniny Mts. B. parvulus was found in the same moss cushions as B. instabilis sp. nov. or B. weglarskae ; similarly, B. parvulus and B. tatrensis co-occur in mosses in the Tatra Mts.







| PCM |
Polish Collection of Microorganisms |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bryodelphax instabilis
| Gąsiorek, Piotr & Degma, Peter 2018 |
Bryodelphax instabilis
| Gąsiorek & Degma 2018 |
Bryodelphax instabilis
| Gąsiorek & Degma 2018 |
Bryodelphax instabilis
| Gąsiorek & Degma 2018 |
Bryodelphax instabilis
| Gąsiorek & Degma 2018 |
Bryodelphax parvuspolaris Kaczmarek et al ., 2012
| Kaczmarek et al. 2012 |
Bryodelphax iohannis Bertolani et al ., 1996
| Bertolani et al. 1996 |
Bryodelphax sinensis (
| Pilato 1974 |
Bryodelphax weglarskae (
| Pilato 1972 |
Bryodelphax tatrensis (Węglarska, 1959)
| Weglarska 1959 |